葉
葉(は、英: leaf[注釈 1])は、陸上植物の植物体を構成する軸性器官である茎に側生する器官である[1]。維管束植物の胞子体においては根および茎とともに基本器官の一つで、シュート頂から外生的に形成される側生器官である[2]。普通、茎に側生する扁平な構造で[2]、維管束からなる脈系を持つ[1]。コケ植物の茎葉体(配偶体)が持つ扁平な構造も葉と呼ばれる[3][1]。

一般的な文脈における「葉」は下に解説する普通葉を指す[4]。葉は発達した同化組織により光合成を行い、活発な物質転換や水分の蒸散などを行う[2]。
葉の起源や形、機能は多様性に富み、古くから葉の定義や茎との関係は議論の的であった[1][2]。ゲーテ以降、葉を抽象的な概念に基づいて定義しようという試みが形態学者によりなされてきたが、ザックス以降、発生過程や生理的機能、物質代謝、そして遺伝子の発現や機能などに解明の重点が置かれている[2]。茎と同様にシュート頂分裂組織に 由来するが、軸状構造で無限成長性を持つ茎とは異なり、葉は一般的に背腹性を示し、有限成長性で腋芽を生じない[2]。維管束植物の茎はほぼ必ず葉を持ち、茎を伸長させる分裂組織は葉の形成も行っているため、葉と茎をまとめてシュートとして扱う[5]。
進化的起源
葉の進化的起源は系統によって異なり、コケ植物の茎葉体(配偶体)が持つ葉 (phyllid)、小葉植物の胞子体が持つ小葉[注釈 2]、そして種子植物の胞子体が持つ大葉は独立に進化してきた[1][3][6]。大葉は形態の変異に富み、針葉などもこれに含まれる[1]。また、大葉植物の内部系統でも、葉は最大で11回独立に進化してきたと考えられている[7]。特に、大葉シダ植物の胞子体が持つ羽葉やトクサ類の楔葉は被子植物の大葉とは異なる起源を持っていると考えられている[8]。大葉シダ植物の中ではマツバラン目では、葉を持たず、茎には葉状突起が側生する[9]。
コケ植物の葉 (phyllid, phyllidium)[10]は配偶体にできる点で大きく異なり、普通1細胞層からなり、維管束がなく中肋という軸で支持され、維管束植物の葉とは起源も形態も本質的に異なるものである[1][11]。
葉の起源を含む包括的な維管束植物の形態進化はヴァルター・マックス・ツィンマーマンが提唱した仮説、テローム説によって解釈される[12][13]。古典形態学の概念では生物がある「原型」を変形させることで進化したと考えらえており、テローム説もその流れに則っている[14]。陸上に進出した当時の陸上植物は二又分枝を行う軸により植物体が構成されていた[12]。ツィンマーマンはそれに基づき、そういった植物は形而上学的な単位である「テローム」及び「メソム」と呼ばれる軸から体が構成されていたと考え、それが癒合や扁平化などの変形をし陸上植物の根や茎や葉を形づくったと考えた[13][14]。二又分枝の末端の枝をテローム、それ以外のテロームを繋ぐ軸をメソムと呼び、二又分枝の体制はそれらの軸を単位として構成されていたとした[14]。
大葉

大葉(だいよう、または大成葉、megaphyll, macrophyll)は葉身に多数の葉脈が形成される葉である[15]。種子植物の大葉と大葉シダ植物の羽葉(うよう、frond)、そして大葉シダ植物のうち基部トクサ類がもつ楔葉(けつよう、sphenophyll、または輪葉[16]、輪生葉[17]とも)が大葉に含まれる[18]。これらの葉はかつては相同であると考えられたこともあったが[18]、現在では何れも進化的起源や性質が異なると考えられている[6]。大葉植物(特に被子植物と大葉シダ植物)の葉跡[注釈 3]の上側の髄と皮層を繋いでいる部分には一次木部細胞に接して柔細胞が形成されている[19]。大葉シダ植物の羽葉では茎から葉原基に向かって葉跡が伸長する[19]。羽葉の葉跡の上にある柔組織を葉隙(ようげき、leaf gap)と呼ぶ[19]。それに対し、被子植物の葉は葉跡が葉原基から茎に向かって伸長する求基的葉である[19][20]。被子植物の葉跡の上にある柔組織は空隙(くうげき、lacuna)と呼ぶ[19]。それぞれの葉の起源も形成過程も異なるため、葉隙と空隙は相同ではないと考えられている[19]。葉隙や空隙の存在は小葉との識別点とされてきたが、葉隙の有無は完全に系統を反映しているわけではない[15]。トクサ類や種子植物の真正中心柱では葉柄に入る葉跡が多数あり、それぞれが茎の維管束から仮軸分枝によって供給されるため葉隙はなく、メシダ科など薄嚢シダ類でも網状中心柱が小型化すると葉跡が仮軸分枝するため、見かけ上葉隙がなくなる[15]。また、トクサ類の楔葉は節に輪生し、小葉のように葉跡は1本であるが、古い時代のものでは脈が又状分岐するのもある[16][21]。構造が単純化した現生のトクサ属のものは葉緑体を持たず光合成は行わないようになっており、葉の基部が隣同士で融合して袴状の葉鞘を作るものがある[16][22]。しかし化石植物の楔葉はそれより大型であり、プセウドボルニア Pseudobornia では2回二又分枝した軸に細かい葉片が鳥の羽状につく形態であった[22]。かつては葉隙の有無に焦点が当てられていたこともあり、葉隙ができないトクサ類の楔葉は小葉であるとされていた[23]。
大葉植物の葉はテローム説における癒合および扁平化により形成されたという解釈がなされている[24][25]。大葉の完成には、テローム軸が癒合および扁平化することに加えて背腹性と左右相称性の獲得が必要であった[26]。現生大葉植物のステム群であるトリメロフィトン類 Trimerophytopsida では、二又分枝の2本の枝に強弱が生じ不等二又分枝を行うか、無限成長をする主軸と側軸の分化が起こり、単軸分枝するようになった[24][25]。また、側軸が平面に展開する傾向がある[25]。この2つの性質は大葉の形成途上と考えることができ[25]、葉の祖先である軸が側生器官の特徴を獲得した段階であると考えられる[26]。軸の癒合による葉面形成はトリメロフィトン類ではまだ進んでおらず、そこから派生した各系統で葉面形成が起こったと考えられている[7]。
テローム説では二又分枝を行っていた植物が持つテローム軸が癒合し、扁平化することで大葉植物が持つ扁平な葉が形成されたと考えられているが、すでに出来上がった枝が癒合することはないため、テローム説を現代的な生物学に対応させて考えれば、複数の器官の集まりである枝系を作っていた発生遺伝子系が1つの器官である葉を作る発生遺伝子系へと進化したと解釈できる[24]。しかし、現生植物の葉でシュート頂分裂組織で機能する遺伝子制御系が機能していても、葉にシュート頂分裂組織の遺伝子系が流用されているだけかもしれないという可能性が否定できず、側枝から葉が進化した証拠としては乏しい[27]。また上記の通り、大葉は多数回起源であり、それぞれの葉形成の仕組みが共通しているとは必ずしも言えない[28]。
中期デボン紀から後期デボン紀にかけての種子植物の祖先における扁平な葉身の獲得は、葉の進化において鍵となるイベントであった[29]。この扁平な葉身は光の捕捉効率を最大化させるとともに、背腹性を獲得し、葉に向軸側と背軸側の2領域を作り出した[29]。向背軸極性を決めるのはYABBY遺伝子群とKANADI遺伝子群である[30]。YABBY遺伝子群は被子植物の葉形成に関わり現生裸子植物でも保存されているが、種子植物以外には存在しない[28][31]。そのため、大葉形成の遺伝子系は種子植物か木質植物の共通祖先でできあがった可能性がある[31]。
大葉シダ植物においては、化石植物群であるコエノプテリス類 Coenopteridales のスタウロプテリス科とジゴプテリス科では茎と羽葉の分化が不十分で、不完全な背腹性を獲得していた[32]。葉柄に当たる部分の維管束はまだ放射相称で葉態枝と呼ばれ、分枝が進んだ頂端付近の羽軸や小羽軸で背腹性が生じる[32]。現在の大葉シダ植物が持つ羽葉では背腹性および左右相称性を獲得している[20]。
小葉

小葉(しょうよう、または小成葉、microphyll)は原生中心柱や板状中心柱から葉隙を形成せず生じ、通常1本のみの葉脈が通る葉である[33][15]。小葉植物の葉の起源は、突起仮説に基づいた解釈が有力だと考えられている[33][13]。ほかにテローム説の1つであるテローム軸の退縮説、胞子嚢を頂生する軸の退化説がある[13][34]。後二者の仮説は証拠に乏しいが、完全に否定されたわけではなく、今後の小葉類の分子発生学的研究による解明が俟たれる[13]。
突起仮説は1935年、フレデリック・バウアーによって提唱されたもので、軸の表面に生じた棘状の突起が進化の過程で大きくなり、そこに維管束が入り込むことによって形成されたとするものである[33][13][34]。これは化石証拠が得られている[13]。すなわち、小葉植物のステム群であるゾステロフィルム類のソードニア Sawdonia では維管束を持たない突起のみが存在し、現生小葉植物の姉妹群であるドレパノフィクス類のアステロキシロン Asteroxylon では維管束は突起の付け根まで伸び、古生リンボク目のレクレルキア Leclercqia や現生小葉植物では小葉中に1本の葉脈がみられる[33][13]。
葉状突起

大葉シダ植物ハナヤスリ亜綱のマツバラン目では、葉を持たず、茎には葉状突起(ようじょうとっき、foliar appendage)が側生する[9][35]。マツバラン属 Psilotum の葉状突起には維管束がないが、イヌナンカクラン属 Tmesipteris の葉状突起は葉隙がなく、1本の維管束が伸びている[9]。また、ソウメンシダ Psilotum complanatum では分枝した維管束が葉状突起の基部まで伸びている[35]。これは小葉植物の小葉と類似しているが、別起源である[9]。
外部形態
葉緑体を持ち、光合成を行う葉を普通葉(ふつうよう、foliage leaf)と呼ぶ[4][36]。普通葉の多くは扁平であるが、針葉樹の針状葉(しんじょうよう、needle leaf)やネギ属 Allium(ヒガンバナ科)やイグサ属 Juncus(イグサ科)が持つ管状葉(かんじょうよう、tubular leaf)も普通葉に含まれる[4]。また、1個体に異なる形態の普通葉が生じる現象を異形葉性(いけいようせい、heterophylly)と呼ぶ[37][注釈 4]。より広義には、普通葉の形態に限らずその種の特徴として常に2種類以上の異なる形態の葉を持つことを指す[37][39]。異形葉性を示す葉を異形葉(いけいよう、heterophyll)という[39]。
葉の構成部分は基部から順に、托葉、葉柄、葉身の3部に大別される[4][36]。托葉(たくよう、stipule)は葉の基部付近の茎または葉柄上に生じる葉身とは異なる葉的な器官で[4]、葉柄(ようへい、petiole)は茎と葉身を繋ぎ、葉身を支持する[40]。被子植物の葉が持ち、普通扁平な光合成を行う主要な部分を葉身(ようしん、lamina, blade)という[29][40]。葉身の組織は葉脈、葉肉、表皮からなる[40]。
托葉や葉柄を欠く葉も多い[4][40][41]。葉柄を欠く葉を無柄葉(むへいよう、sessile leaf)という[40]。また、葉身を欠くものもあり、偽葉(ぎよう、phyllode)と呼ばれる[41][42]。
普通葉の形状から木本植物を大別した場合、広葉樹(こうようじゅ、broad-leaved tree, hardwood)と針葉樹(しんようじゅ、needle-leaved tree, acicular tree)に分けられる[43]。基本的には系統関係と対応しているため、イチョウ Ginkgo biloba(イチョウ科)、ソテツ Cycas revoluta(ソテツ科)、ナギ Nageia nagi およびイヌマキ Podocarpus macrophyllus(マキ科)といった裸子植物は広葉をもつが広葉樹ではない[43]。このうち、マキやナギは、鱗状葉を持つヒノキやイブキ(ヒノキ科)、針状葉を持つマツ科や旧スギ科とともに針葉樹に含まれる[43]。ガンコウランやツガザクラなどの針状の葉(エリカ葉)を持つ広葉樹もある[43][44]。イチョウやソテツ、ヤシ類はどちらにも含まれない[43]。また、針葉樹の葉は形態によって針形葉、線形葉、鱗形葉に分けられる[45](下記「#針葉樹の普通葉」節を参照)。
複葉
%252Clf%252Ce-s_greenhouse_Radix_Serre%2540Wageningen_University%252CNL_fri24jun2011.jpg.webp)
葉身が複数の小部分に分かれた葉のことを複葉(ふくよう、compound leaf)とよぶ。それに対し、それに対して、葉身が1枚の連続した面からなる葉を単葉(たんよう、simple leaf)と呼ぶ[46]。複葉は単葉の葉身の切れ込みが深くなり、主脈の部分にまで達した状態であると解釈される[47]。
複葉における、分かれている葉身の各片を小葉(しょうよう、leaflet)、小葉が付着する中央の軸部を葉軸(ようじく、rachis)と呼ぶ[47][48]。小葉が柄を介して葉軸につく場合、その柄は小葉柄(しょうようへい、petiolule)と呼ばれる[47][48]。葉片が単葉か複葉の一部かは腋芽の有無によって区別され、複葉の小葉柄の基部には腋芽ができない[48]。
葉縁の形質
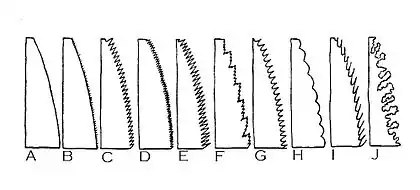
A 全縁、B 毛縁、C–E 鋸歯縁、F 重鋸歯縁、G 歯牙縁、H 円鋸歯状縁、I 微突形、J 条裂
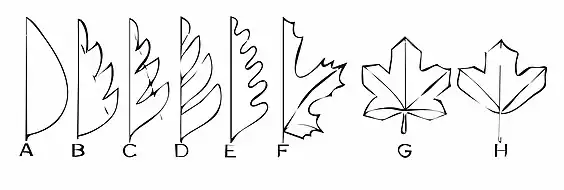
A 全縁の不分裂葉、B 浅裂、C 深裂、D 全裂、E 波状縁、F 欠刻縁、G 掌状葉、H 三裂葉
葉縁にみられる鋸の歯のような細かな切れ込みを鋸歯(きょし、serration, teath)という[50]。鋸歯を持たず、切れ込みもないことを全縁(ぜんえん、entire)という[50][51][52]。
凹凸が大きく葉全体の形にかかわるほどの切れ込みがある単葉を分裂葉(ぶんれつよう、lobed leaf)と呼ぶ[48]。この突出部を裂片(れっぺん、lobe)という[51]。それに対して裂片のない葉を不分裂葉という[53]。切れ込みが浅いものを浅裂(せんれつ、lobed, lobate)、やや深く切れ込むものを中裂(ちゅうれつ、cleft)深く裂けていれば深裂(しんれつ、parted, partile)、完全に裂けたものを全裂(ぜんれつ、dissected)という[51]。裂片が放射状に配置し、掌のようになったものを掌状(しょうじょう、palmate)、裂片が左右に列をなし、鳥の羽のようになったものを羽状(うじょう、pinnate)という[51]。裂ける深さと形を組み合わせて、葉の形状を表現することが多く、例えばヤツデの葉は掌状深裂、ヨモギの葉は羽状深裂する。
有鞘葉

単子葉植物の葉の多くは有鞘葉(ゆうしょうよう、sheathing leaf)となるものが多い[54]。有鞘葉は扁平な部分と基部の葉鞘(ようしょう、leaf sheath)からなる[54]。葉鞘はイネ科、カヤツリグサ科、ツユクサ科、ショウガ科、ラン科などに一般的で、ユリ科の一部にも見られる[54]。
葉鞘はつねに地上茎の節から生じるわけではなく、地下茎から直接生じて順次内側の葉鞘を包み、筒状となって地上茎のように見えることがある[54]。こうした葉鞘の集まりを偽茎(ぎけい、pseiudostem)と呼ぶ[54]。ガマ科、ショウガ科、テンナンショウ属 Arisaema(サトイモ科)、シュロソウ属 Veratrum(シュロソウ科)、スズラン属 Convallaria(キジカクシ科)などに見られる[54]。
葉身が発達せず、葉鞘だけの葉を鞘葉(しょうよう、sheath leaf)と呼ぶ[54]。鞘葉はイグサ科のイグサ Juncus decipiens やミヤマイ Juncus beringensis、カヤツリグサ科のワタスゲ Eriophorum vaginatum やホタルイ Schoenoplectiella hotarui、カンガレイ Schoenoplectiella triangulatus、フトイ Schoenoplectus tabernaemontani、ハリイ属 Eleocharis などに見られる[54]。これらでは稈の基部に小数個の鞘葉が重なり合っている[54]。また、ホシクサ属 Eriocaulon(ホシクサ科)では茎の下部に常に1個の鞘葉がある[54]。
また、有鞘葉のうち花序に腋生するものを苞鞘(ほうしょう、bract sheath)という[55]。スゲ属 Carex の苞は苞鞘であることも無鞘であることもあり、シバスゲ節 sect. Praecoces やシオクグ節 sect. Paludosae の小穂の苞は少なくとも最下が苞鞘である[55]。
根生葉
根生葉(こんせいよう、または根出葉、radical leaf)は地上茎の基部の節に付き、根から生じているように見える葉である[56]。大葉シダ植物や草本性被子植物に多い[56]。バラの花冠状に放射状に重なり合ってつき、地表に密着して越冬する根生葉をロゼット葉(ロゼットよう、rosette leaf)と呼ぶ[56]。
なお、根生葉に対し伸長した地上茎に側生する葉は茎生葉(けいせいよう、または茎葉、cauline leaf)と呼ぶ[56]。
楯状葉
葉柄の先に雨傘状の葉身を持つ葉を楯状葉(盾状葉、じゅんじょうよう、peltate leaf)という[57][58]。ハスやジュンサイ、ノウゼンハレン、サンカヨウ属、ミヤオソウ属、テンジクアオイ属、ハスノハカズラ属などで見られるほか、ヤブレガサやタイミンガサのように葉身が放射状に分裂しているものもある[57][58]。また、楯状葉葉身の葉縁の拡大があまり進行せず、葉身の葉縁方向への平面成長が進んだ形態は、杯状葉または嚢状葉と呼ばれる[59]。杯状葉(盃状葉、はいじょうよう、aecidial leaf)は奇形として知られており、ラッパイチョウやクロトン、シナガワハギなどによく観察されている[59][60][58]。
針葉樹の普通葉

古くから針葉樹類と言われた裸子植物の系統は[61]、分子系統解析が進んだ現在ではマツ科と残りの針葉樹類(広義のヒノキ目)の2系統が含まれることが分かっている[62][63]。現生針葉樹類の普通葉は全て単葉である[62][64]。その中でも、多くの針葉樹類の葉は細くて先細りとなるため、針葉(しんよう、needles)と表現される[64]。ただし、ナギモドキ属 Agathis やナンヨウスギ属 Araucaria(ナンヨウスギ科)、マキ科(ナギ属 Nageia)では著しく幅の広い葉を持つ[64][65]。ヒノキ科以外の多くの針葉樹類の葉は長枝に発生し、螺旋葉序または互生葉序となる[64]。ヒノキ科では全て十字対生葉序か輪生葉序である[64]。
現生針葉樹の葉は、その形態によって針形葉、線形葉、鱗形葉と呼び分けられる[45]。Laubenfels (1953) は現生針葉樹類の葉を、その3つにナギなどの幅広い葉を加えた4つのタイプに分類した[65]。
針状で扁平ではないものを針形葉(しんけいよう、または針状葉、針葉、needle leaf)という[45][4][44]。スギは針形葉が螺旋状につき、葉の基部が小枝と一体化している[45]。マツ属 Pinus ではシュートに長枝と短枝が分化し、針形葉が短枝に分類群ごとに1–5本の一定の数ずつつく[45][66][64]。この短枝は俗に「松葉」と呼ばれる[66]。クロマツでは短枝に2本の針形葉、ダイオウマツは短枝に3本の針形葉、ゴヨウマツは短枝に5本の針形葉をつける[45]。また、マツの葉は等面葉である[44]。
幅が狭く扁平なものを線形葉(せんけいよう、または線状葉、線葉)という[67]。中脈が明らかで、背軸面には気孔が気孔帯がみられることが多い[67]。モミ、ツガ(マツ科)、カヤ、イヌガヤ(イチイ科)などには2本の気孔帯が認められる[67]。イヌマキ(マキ科)の線形葉は中脈が顕著である[67]。コウヤマキ(コウヤマキ科)の線形葉は短枝につく2本の葉が合着したものである[67]。
扁平な葉が十字対生して茎を包んでいるものを鱗形葉(りんけいよう、または鱗状葉、鱗葉、scale like leaf)と呼ぶ[68][69][注釈 5]。ヒノキ科の普通葉に多く[69]、ヒノキやサワラ、アスナロやコノテガシワに見られる[68]。ビャクシンの葉は普通、鱗形葉であるが、ときどき針形葉を交じる[68]。
葉の特殊化
葉は地上の茎に付属し、扁平で光合成を行うのが典型であるが、付く位置や形、機能においてさまざまな特殊化がみられる[56]。こうした葉と相同と考えられるものの光合成を担うわけではない器官と普通葉とを合わせて総称的に葉的器官(ようてききかん、phyllome, foliar appendage、フィロム[29])と呼ぶこともある[44]。葉的器官には普通葉や芽鱗、苞、花器官などが含まれる[29]。
鱗片葉

普通光合成を行わず、普通葉に比べ著しく小型化した葉を鱗片葉(りんぺんよう、scale leaf, scaly leaf)と呼ぶ[69]。裸子植物の鱗片葉は雄性胞子嚢穂(雄性球花)、イチイ科の雌性胞子嚢穂(雌性球花)、マツ科の長枝等にみられる[69]。
鱗片葉はさらに特殊化し、その位置により様々に呼び分けられる[69]。芽を覆う鱗片葉は芽鱗(がりん、bud scale)、花芽を腋にもつ鱗片葉は苞(ほう、または苞葉、bract)と呼ばれる[69]。苞は位置や形により、総苞、苞、小苞、苞鞘、苞穎などに分けられる[70]。
生殖シュートにおいて、胞子嚢とそれに由来する構造以外の要素は葉に由来すると考えられている[71]。萼片、花弁、雄蕊、心皮といった被子植物の花を構成する鱗片葉を花葉(かよう、floral leaf)[71][69][72][73]または花器官(はなきかん、floral organ)という[29]。雄蕊や雌蕊は胞子葉(ほうしよう、sporophyll)が変形してできたものである[74]。また、裸子植物の雌性胞子囊穂(雌性球花、球果)を構成する鱗片葉は種鱗(しゅりん、ovuliferous scale, seed scale)と苞鱗(ほうりん、bract scale)の2種類からなり、それらが癒合して種鱗複合体(しゅりんふくごうたい、seed scale complex)を構成する[69][75][76]。種鱗複合体は果鱗(かりん、fructiferous scale, cone scale)や苞鱗種鱗複合体とも呼ばれる[69][75]。
シュートの下部に形成される鱗片葉は低出葉(ていしゅつよう、cataphyll、独: Niederblätter)と呼ばれる[42][77]。低出葉には鞘葉、芽鱗、芽鱗に似た托葉だけの葉、実生の上胚軸の下部に作られる鱗片葉などがある[42]。鞘葉は単子葉類の茎の下部にみられる[42]。芽鱗は鱗芽をもつ木本に普通にみられる[42]。托葉だけの葉はキジムシロ属のキジムシロ Potentilla fragarioides、イワキンバイ Potentilla ancistrifolia var. dickinsii、ミツモトソウ Potentilla cryptotaeniae などにみられる[42]。クスノキ科のタブノキ属 Machilus、クロモジ属 Lindera[注釈 6]などの実生では、子葉の間から伸びた上胚軸が地上に出ると互生する鱗片葉を形成する[42]。この鱗片葉は次第に普通葉へ移行する[42]。多くの被子植物では、シュート発生の際に周期的に低出葉の形成が起こる[77]。また、こうして作られた側枝の最下の低出葉は前出葉(ぜんしゅつよう、prophyll, fore-leaf)と呼ばれる[42]。前出葉は側芽に最初に作られ、特殊な形態を示すことが多い[42]。ミカン属 Citrus の葉腋に出る刺やイネ科の小穂の第一苞穎および第二苞頴、スゲ属の果胞および小穂の柄の基部に生じる鞘葉は前出葉である[42]。
シュートの上部に形成される花葉以外の特殊な葉を高出葉(こうしゅつよう、hypsophyll)と呼ぶ[42]。高出葉は狭義には総苞片、苞、小苞などの鱗片葉が含まれるほか、広義にはシュートの上部にあって変質や退化した葉も含まれる[42]。ウスユキソウ属 Leontopodium(キク科)の頭花群の下に伸びる毛深い苞、トウダイグサ属 Euphorbia(トウダイグサ科)の杯状花序の基部にある対生葉、ネコノメソウ属 Chrysoplenium(ユキノシタ科)の花序に含まれる苞以外の黄色い部分などがその例である[42]。
エリカ葉

エリカ葉(エリカよう、ericoid leaf)はツツジ科のガンコウラン属 Empetrum やツガザクラ属 Phyllodoce、エリカ属 Erica などが持つ小さく針状の葉で、重複葉(ちょうふくよう、duplicate leaf)とも呼ばれる[69][44][78]。葉縁付近の背軸側(腹側)に襞状の突起ができ、葉の背軸側に空洞部分ができることで気孔をその空洞の内側にのみ持つようになっている[44]。左右の葉縁が背軸側に折れ曲がったように見えるが、実際は発生の途上に背軸側の基本組織中に新たに生じた分裂組織から二次的に作られたものである[69]。この部分を重複葉身(ちょうふくようしん、duplicate blade)という[69]。気孔が分布する空洞に面していない部分は厚いクチクラに覆われ、クチクラ蒸散を極度に減らしている[79]。また、気孔の分布する空洞部分と外界を連絡する溝の両側は毛が覆い、空気の流通を妨げている[79]。逆に葉の向軸側の表皮下には日射の強い高山において光合成効率を上げるため柵状組織が発達している[79]。こうした構造により蒸散を最小限に抑え[80]、高山に適応している[44]。
水生植物の葉

.jpg.webp)

水生植物の葉は水辺環境に適応して特殊化しており、水面との位置関係により沈水葉、浮水葉、抽水葉が区別される[81]。また、ホテイアオイ Eichhornia crassipes(ミズアオイ科)などの浮遊植物では浮き袋(うきぶくろ、air bladder)を持つ[81]。
- 沈水葉
沈水葉(ちんすいよう、submerged leaf)は、水中にある沈水性(ちんすいせい、submergence)を持つ葉である[81]。一般に軟弱で、機械的組織の発達が悪い[81]。バイカモ Ranunculus nipponicus var. submersus(キンポウゲ科)、マツモ Ceratophyllum demersum(マツモ科)、タヌキモ Utricularia vulgaris(タヌキモ科)、クロモ Hydrilla verticillataやセキショウモ Vallisneria asiatica(トチカガミ科)、エビモ Potamogeton crispus(ヒルムシロ科)など見られ、これらは全ての葉が沈水性を持つ[81]。
- 浮水葉
浮水葉(ふすいよう、または浮葉、floating leaf)は、水面に浮かぶ浮水性(ふすいせい、floatage)を持つ葉である[81]。気孔は水面と反対の向軸面にある[81]。デンジソウ Marsilea quadrifolia(大葉シダ植物デンジソウ科)、ヒツジグサ Nymphaea tetragona(スイレン科)、ジュンサイ Brasenia schreberi(ハゴロモモ科)、ヒシ Trapa jeholensis(ミソハギ科)、トチカガミ Hydrocharis dubia(トチカガミ科)、ヒルムシロ Potamogeton distinctus(ヒルムシロ科)、アサザ Nymphoides peltata やガガブタ Nymphoides indica(ミツガシワ科)などが持つが、若い葉では沈水性を持つことが殆どである[81]。イチョウバイカモ Ranunculus nipponicus (キンポウゲ科)は多くが沈水葉だが、僅かに水面上か水中にある扇形の浮水葉を持つ[81]。
- 抽水葉
抽水葉(ちゅうすいよう、または挺水葉、emergent leaf)は、水面に抜き出る抽水性(ちゅうすいせい、emergence)を持つ葉である[81]。浅水域に生える、ハス Nelumbo nucifera(ハス科)、コウホネ Nuphar japonica(スイレン科)、オモダカ Sagittaria trifolia やクワイ Sagittaria trifolia 'Caerulea'(オモダカ科)、ガマ Typha latifolia(ガマ科)などが持つ[81]。ハスやコウホネは若い葉は浮水性を持つ[81]。
- 根葉
水生シダ類のサンショウモ Salvinia natans の葉は異形葉性を示し、水面に浮かぶ2枚の浮葉(気葉)のほかに、水中に分枝した根状の根葉(こんよう、root leaf)を持つ[82][83]。
貯蔵葉
.JPG.webp)
柔細胞が多量の貯蔵物質を具え、多肉質になった葉を貯蔵葉(ちょぞうよう、storage leaf)という[70]。ユリ属 Lilium やネギ属 Allium の鱗茎(地下茎)は肥厚した貯蔵葉が集合してでき、鱗茎葉(りんけいよう、bulb leaf)と呼ばれる[70][84]。クロユリ Fritillaria camtschatcensis(ユリ科)のもつ鱗茎葉は米粒から豆粒大の立体形をしている[70]。
捕虫葉
食虫植物が持つ、昆虫などの動物を捕らえるように変形した葉を捕虫葉(ほちゅうよう、insectivorous leaf)という[70]。捕虫葉の形は様々で、様々な捕虫の方法がある[70]。モウセンゴケ類 Drosera の捕虫葉は葉縁や葉の表面に長い腺毛を持ち、触れると粘液を出して葉身を巻き込み虫を捕まえる[70]。ムシトリスミレ Pinguicula vulgaris やコウシンソウ Pinguicula ramosa(タヌキモ科)の捕虫葉は表面に腺毛と無柄の腺が密生し、前者からは粘液、無柄腺からは消化液を分泌し、虫を捕らえる[70]。
捕虫葉が嚢状に変化して、捕虫嚢(ほちゅうのう、insectivorous sac)を形成するものもある[70]。嚢状葉(のうじょうよう、pitcher)とも呼ばれる[80][85][83]。タヌキモ属の葉は葉身が小さな捕虫嚢となっており、内部を減圧することで虫を吸い込む[70]。ウツボカズラ属 Nepenthes の葉は葉の先が葉巻きひげとなり、その先が捕虫嚢となっている[70]。サラセニア属では葉柄が漏斗状の捕虫嚢となっている[70]。特にムラサキヘイシソウ Sarracenia purpurea では、その形成過程が明らかになっている[86]。シロイヌナズナのような平面葉と同様に向背軸を規定する遺伝子が発現するが、葉の基部側の細胞分裂の方向が変化することにより、嚢状葉が形成される[86]。ウツボカズラ属やサラセニア属の捕虫嚢内部には毛が生えて虫の脱出を防いでいる[70]。
葉巻きひげ

植物が持つ巻きひげのうち、托葉や葉柄、小葉や葉身の一部を変形させてできたものを葉巻きひげ(はまきひげ、または葉性巻きひげ[80]、leaf tendril)という[70]。バイモ Fritillaria verticillata(ユリ科)の上部の葉の先や葉全体が、トウツルモドキ Flagellaria indica(トウツルモドキ科)では葉の先が巻きひげとなる[70]。マメ科のソラマメ属やレンリソウ属では頂小葉が巻きひげに置き換わった羽状複葉である巻きひげ羽状複葉を形成する[87]。シオデ属 Smilax(サルトリイバラ科)では托葉、ボタンヅル Clematis apiifolia では葉柄、カザグルマ Clematis patens(ともにキンポウゲ科)では小葉柄が巻きひげとなる[70]。なお、葉巻きひげに対し、葉ではなく茎が変形してできた巻きひげになったものは茎巻きひげと呼ばれる[88]。
葉針
葉針(ようしん、leaf spine/needle/thorn)は、葉全体または複葉の小葉、托葉などが硬化して鋭い突起に変形したものである[81]。光合成の機能を持たない[81]。特に托葉が変化した葉針を托葉針(たくようしん、stipular spine)という[81]。葉針に対し、茎が変化したものは茎針[81]、根が変化したものは根針といい、相似器官である[89]。
サボテン(サボテン科)の刺は葉針の一種である[81]。また、メギ Berberis thunbergii やヘビノボラズ Berberis sieboldii(メギ科)では、長枝上に単一または三岐した葉針を生じ、その腋に短枝を形成し、普通葉をつける[81]。ニセアカシア Robinia pseudoacacia(マメ科)は托葉針を持つ[81]。
内部形態
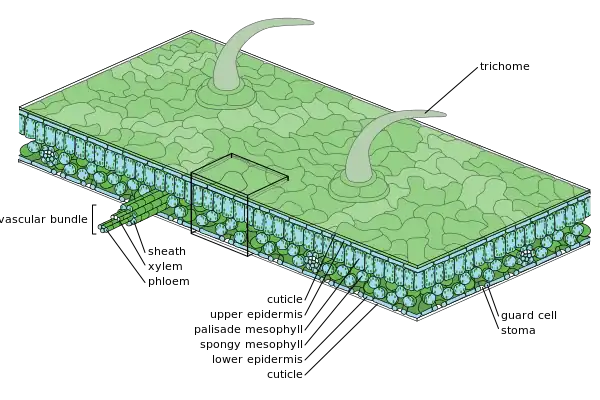
棒状の概形で放射状の構造を持つ根や茎と異なり、葉は左右相称で、背腹性を持つ[90][91]。上側は向軸面、下側は背軸面と呼ばれる[90]。
葉の組織系はザックスの分類 (1875) に基づき、表皮系、基本組織系、維管束系の3つに分けられる[2][91][92][93]。
表皮系
表皮系(ひょうひけい、epidermal system)は表皮細胞、気孔や水孔を作る孔辺細胞、毛状突起(毛、鱗片など)などの構造からなる[91][2]。表皮系は前表皮に由来する[94]。
植物体の表面はふつう1層の表皮細胞からなる表皮(ひょうひ、epidermis)で覆われる[91][94]。ただし複数の細胞層からなる表皮もあり、多層表皮(たそうひょうひ、multiseriate epidermis)と呼ばれる[91]。表皮細胞の外壁には長鎖脂肪酸または蝋を主成分とするクチクラ(cuticule)が分泌されクチクラ層(cuticular layer)を形成することで体表からの水分蒸散を防いでいる[91][95]。クチクラを構成する脂質は陸上植物の中で多様性がある[95]。コケ植物の配偶体および胞子体、小葉植物と大葉シダ植物の配偶体ではクチクラは発達しない[95]。被子植物でも、乾燥地域に生育する植物ではクチクラの発達がよい[95]。維管束植物のクチクラには疎水性細胞外生体高分子であるクチンが含まれている[95]。コケ植物のクチクラにはスベリン様の疎水性細胞外生体高分子を持つ[95]。
気孔(きこう、stoma)は2つの孔辺細胞に囲まれた小間隙で、光合成や呼吸、蒸散などのガス交換のための空気や水蒸気の通路である[91]。
基本組織系
葉の基本組織系は葉肉(ようにく、mesophyll)と呼ばれ[2]、上下両表皮間に挟まれた柔組織からなる[96][97]。葉緑体に富み、同化やガス交換に適した組織への分化が起こっている[2][93]。葉肉は普通葉では同化組織、貯蔵葉では貯蔵組織や貯水組織からなり、鱗片葉ではほとんど発達しない[98][96]。
被子植物の典型的な普通葉では葉肉は向軸側が柵状組織、背軸側が海綿状組織に分化する[96]。柵状組織(さくじょうそしき、palisade tissue)は向軸側にあり、葉面に垂直な方向に比較的密に並んだ細胞からなる[98]。この表皮直下に1層から数細胞層を構成する細胞を柵状柔細胞(さくじょうじゅうさいぼう、palisade parenchyma cell)という[97]。海綿状組織(かいめんじょうそしき、spongy tissue)は背軸側にあり、形や並び方が不規則で、細胞間隙に富んだ組織である[98][99]。これを構成する細胞を海綿状柔細胞(かいめんじょうじゅうさいぼう、spongy parenchyma cell)といい、柵状柔細胞から背軸側表皮の間を埋めている[97]。柵状組織の厚さは陰葉より陽葉でよく発達する[99]。
向軸面に柵状柔細胞、背軸面に海綿状柔細胞が分布し、背腹性がある普通葉を両面葉(りょうめんよう、bifacial leaf)という[54][100]。ネギ属 Allium(ヒガンバナ科)やアヤメ属 Iris(アヤメ科)の葉は円筒形または二つ折れとなり、外観では背軸側のみが見える単面葉(たんめんよう、unifacial leaf)と呼ばれる[54][100]。アヤメ属は両面の表皮下に柵状組織、海綿状組織がある[101]。スイセン属 Narcissus(ヒガンバナ科)では上下表皮下に柵状組織、中央に海綿状組織がある[101]。また、マツ科の針状葉は維管束の特徴で背腹性が分かるが、外観では区別ができないようになっており、等面葉(とうめんよう、equifacial leaf)と呼ばれる[100]。針葉樹類やイネ科の葉は柔細胞が葉肉中にほぼ均等に分布する[101]。
表皮下にある、葉肉の最外層の1から数細胞層の組織を下皮(かひ、hypodermis)という[101]。下皮は葉緑体を持たず、多層表皮の内側の層に似ているが、発生学上表皮と異なり、葉肉と同一の起源を持つ[101]。針葉樹類の下皮は、多くは1–2層の繊維状の厚壁細胞からなる[101]。マツ属 Pinus(マツ科)、スギ Cryptomeria japonica(ヒノキ科)、コウヤマキ Sciadopitys verticillata(コウヤマキ科)では気孔を除いた全周にあるが、ツガ属 Tsuga では葉の両縁部分にのみ見られる[101]。イチイ科にはない[101]。被子植物は下皮を持たないことが多いが、モチノキ属 Ilex(モチノキ科)では背軸面表皮の下に内側の葉肉細胞より少し大きな厚壁細胞からなる下皮を持つ[101]。
葉肉の最内層にあり、維管束を囲む厚壁細胞あるいは柔細胞からなる1層の表皮状の細胞層を内皮(ないひ、endodermis)という[101]。大葉シダ植物や裸子植物の葉には内皮があるが、被子植物にはない[101]。また、針葉樹類の針葉には、内皮と維管束の間に柔細胞と仮道管が入り混じった移入組織(いにゅうそしき、transfusion tissue)がある[101]。移入組織は維管束と葉肉を連絡する補助的な通道組織であると考えられる[101]。
維管束系
葉の維管束系(いかんそくけい、vascular syetem)は葉脈(ようみゃく、vein, nerve)と呼ばれる[2][93]。葉脈は茎の維管束と接続し、その部分を葉跡(ようせき、foliar trace, leaf trace)という[2]。葉肉内における葉脈の分岐の仕方を脈系(みゃくけい、venation)といい、脈系は系統によって多様である[2]。
C4植物の葉
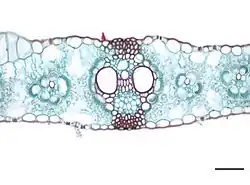
C4植物の葉には、維管束鞘が2重となっており、メストム鞘(メストムしょう、mestome sheath)と呼ばれる[102]。これを欠くC4植物もある[79]。その外側には比較的大きな柔細胞からなる環状葉肉(かんじょうようにく、kranz)がある[102]。こちらは必ず存在し、葉緑体に富んでいる[79]。維管束の外側を維管束鞘が、その外側を葉肉細胞が放射状に取り囲むこの構造を、ドイツ語の「花環 Kranz」からクランツ構造(クランツこうぞう、Kranz anatomy)という[103]。以外の葉肉細胞では柵状柔細胞と海綿状柔細胞の区別が不明瞭である[79]。また、葉脈間の距離がC3植物に比べて短く、空気間隙も少ない[79]。
発生

シュート頂において葉原基(ようげんき、leaf primordia)として外生的に形成され、その発生位置によって葉の配列様式(葉序)が決定する[2][104]。葉原基から葉身・葉柄・托葉が分化し、同時に表皮系・基本組織系・維管束系の組織分化が進行する[2]。
葉原基の形成
葉は、まずシュート頂分裂組織(茎頂分裂組織、SAM)の側方に葉原基として形成される[105]。種子植物のシュート頂分裂組織の細胞は外側からL1、L2、L3の3層の異なる安定的な組織層として組織化されている[105]。葉原基形成にはL1層でのオーキシン極性輸送が必須である[105]。多くの被子植物では、葉原基はシュート頂側面の表面付近の1層から数細胞層の並層分裂に由来する[104]。特に真正双子葉類では通常L2の細胞に最初の並層分裂がみられるが、イネ科などでは外側の2層の細胞分裂に由来する[104]。
1つの葉原基が発生してから次の葉原基が発生するまでの時間を葉間期(ようかんき、plastochron、プラストクロン)という[106]。対生葉序では葉原基が同時に2個形成されるため、次の1対が作られるまでの時間を葉間期とする[106]。シュート頂分裂組織から葉原基が突起すると茎頂は最小の大きさとなり、このときを最小期(さいしょうき、minimal area phase)という[107]。逆に葉原基が分離する直前の茎頂は最大の大きさになり、このときを最大期(さいだいき、maximal area phase)という[107]。ヒョウタンボク属では葉間期は1.5–5.5日であることが分かっている[107]。
葉面の成長
葉原基ははじめ頂端分裂組織を形成し先端成長を始めるが、大葉シダ植物以外ではすぐにその活動が衰退する[104][108]。次に葉原基で向背軸が決定され、それぞれの側で発現する遺伝子が互いに両者を抑制しあうことによって形成される[86]。この過程に働く遺伝子群について、1型YABBY遺伝子群の働きでシュート頂分裂の制御系が抑制され、葉のアイデンティティが付与される一方、HD-ZIPⅢ遺伝子群やKANADI遺伝子群の働きによって背腹性が確立する[104]。続いて、向軸側と背軸側両方の遺伝子の制御によって葉縁部で細胞分裂活性が高くなる[86]。それにより、向軸側と背軸側の境界部分が細胞成長し、扁平な葉面が成長する[86]。
複葉や楯状葉の形成
複葉原基では、本来シュート頂分裂組織で発現し葉原基では発現しない1型KNOX遺伝子[注釈 7]や葉原基とシュート頂の境界で発現し1型KNOX遺伝子の発現境界を規定するCUP-SHAPED COTYLEDON遺伝子(CUC)の発現がみられる[109]。葉原基基部の周縁部 (marginal blastozone) にて1型KNOX遺伝子などの働きにより小葉原基が生じ、葉形が複雑化する[104]。
楯状葉では、裏側を規定する遺伝子が葉原基の基部では葉の表側に発現していることで細胞分裂活性の高い領域が円形になり、形成されると推定されている[86]。
大葉シダ植物の葉
また、被子植物の成熟した葉は分裂組織や幹細胞を持たないが、小葉植物のイワヒバ科、大葉シダ植物のトクサ類と薄嚢シダ類の葉の頂端には2面切り出しの幹細胞が存在する[110][111]。また、典型的な薄嚢シダの葉縁にある周縁分裂組織は4面切り出し、コケシノブ科の葉縁では1面切り出しの幹細胞が存在する[110]。
1つの腋芽とセットとなって1つの単位を形成し、成長と組織形成が求基的に進む被子植物の葉と異なり、大葉シダ植物の羽葉ではワラビ巻きから生じて求頂的に成長する[20][111]。薄囊シダ類の葉の頂端にはレンズ型(3面体)の頂端幹細胞があり、全ての葉細胞の母細胞となる[111][112]。また、葉縁にも幹細胞を持つ周縁分裂組織(しゅうえんぶんれつそしき、marginal meristem)が形成される[111][112]。特にウラジロ科のウラジロやコシダ、カニクサ科のカニクサ、コバノイシカグマ科のワラビやユノミネシダなどでは数年に亘って頂端幹細胞が分裂を続け、葉の先端部分が無限成長して羽片を作り続けることから、種子植物より茎的な性質を保持している[113][27]。
コケ植物の葉
被子植物とは異なる発生機構によって形成されている[86]。
蘚類、特にモデル植物であるヒメツリガネゴケ Physcomitrella patens(ヒョウタンゴケ科)の茎葉体は、配偶体の別のステージであるカウロネマ細胞が形成した側枝始原細胞から、約5%の確率でオーキシンの作用により転写因子ABPが誘導され、茎葉体頂端幹細胞になることで形成される[3]。茎葉体頂端幹細胞から切り出された細胞はセグメント細胞と呼ばれ、並層分裂を行って先端側と基部側の2つの娘細胞を形成する[114]。そのうち先端側の細胞が垂層分裂を行い、形成された茎葉体頂端幹細胞に近い方の細胞が葉頂端幹細胞となる[114]。葉頂端幹細胞は2面切り出しの頂端幹細胞で[110]、1枚の全ての葉を形成する[114]。
胚発生
種子植物胞子体の個体発生において、最初に形成される葉を子葉(しよう、cotyledon)という[115][116]。かつての植物分類体系では、子葉の枚数に基づいて被子植物を子葉が2枚の双子葉類と子葉が1枚の単子葉類に分類してきたが、分子系統解析により双子葉は共有原始形質であり、系統的には正しくないことが分かっている[115]。双子葉植物の子葉は対生し、ふつう同形で主軸の子葉節につく[116]。
園芸界では、双子葉植物の実生において、展開した地上生子葉または地表性子葉を双葉(ふたば)といい、それに対して普通葉を本葉(ほんば)という[117]。
葉序
葉序(ようじょ、phyllotaxis)は、茎に対する葉の配列様式である[55][118]。葉序は節につく葉の枚数により、1節に1枚葉がつく互生葉序と1節に2個以上の葉がつく輪生葉序に分けられる[55]。輪生葉序のうち、1節に2個ちょうどの葉をつける葉序を特に対生葉序と呼び分けることも多く、葉序は普通、互生葉序(ごせいようじょ、alternate phyllotaxis)、対生葉序(たいせいようじょ、opposite phyllotaxis)、輪生葉序(verticillate phyllotaxis)の3つに大別される[55][118]。
葉原基形成の際の、シュート頂分裂組織におけるL1層でのオーキシン極性輸送が葉序を生み出す要因となっている[105]。
葉上生

葉は有限成長する側生器官であるため、他の器官を付けないのが普通であるが、葉上に花序や不定芽を付けることがあり、このような性質を葉上生(ようじょうせい、葉上形成、epiphylly)という[37][119]。
ハナイカダ属 Helwingia(ハナイカダ科)やビャクブ Stemona japonica(ビャクブ科)は普通葉と花序が発生初期に原基が分かれることなく同時に成長し、葉上に花序ができる[37]。シナノキ属 Tilia(アオイ科)では苞上に花序が生じるように見える[37]。芽の下側にあって脇芽を戴く葉を蓋葉(がいよう、subtending leaf)といい、これらは蓋葉と脇芽が癒合してできたものである[120][121]。そのため、これは不定芽とは異なる[121]。
葉上不定芽
葉上芽(ようじょうが、または葉上不定芽、epiphyllous bud)は脱分化により葉に生じた不定芽である[122][123][121]。
コダカラベンケイ Kalanchoe daigremontiana やセイロンベンケイ Kalanchoe pinnata、キンチョウ Kalanchoe tubiflora、Kalanchoe scandens(ベンケイソウ科)などでは普通葉の葉縁に不定芽が生じる[37][124]。この不定芽は受精卵と同様な形態的変化の過程をとって体細胞から生じる不定胚を経て形成される[122][125]。また、Begonia phyllomaniaca も自然状態で葉の表面脈上に無数の不定芽や葉片状形成物を生じる[126]。
単子葉類にも葉上不定芽の例は多く知られ、ショウジョウバカマ Heloniopsis orientalis(シュロソウ科)は葉の先端付近に不定芽を生じる[122][126]。ショウジョウバカマの不定芽は中央脈維管束の木部に近い数個の細胞が分裂し、カルス状の隆起から内生的に形成される[126]。ヤチラン Malaxis paludosa は葉の先端付近の葉縁に不定芽を生じ、脱落して別個体となる[126]。カラスビシャク Pinellia ternata(サトイモ科)も葉身の基部や葉鞘頂端部に、スルガテンナンショウ Arisaema yamatense では葉鞘頂端部に、離脱しない多肉質の珠芽を生じる[119][122]。
薄嚢シダ類でも多数、葉上不定芽を生じる例が知られている[122][123][127]。クモノスシダ Asplenium ruprechtii やヒメイワトラノオ Asplenium capillipes、チャセンシダ、ヌリトラノオ、ヒノキシダ(チャセンシダ科)、フジシダ Monachosorum maximowicziiやオオフジシダ Monachosorum nipponicum(コバノイシカグマ科)、ツルデンダ Polystichum craspedosorum(オシダ科)では葉の先端に近い表面から不定芽を生じ、独立して新たな個体となる[122][127]。ミズワラビ Ceratopteris thalictroides(イノモトソウ科)は葉縁に、コモチシダ Woodwardia orientalis(シシガシラ科)は葉の表面脈上に多数の不定芽を生じる[128]。これらは葉縁に残存分裂組織 (residual meristem) があるためであると解されている[128]。普通向軸面からのみ不定芽が生じるが、Tectaria cicutaria[注釈 8](ナナバケシダ科)やチリメンシダ Dryopteris erythrosora f. prolifica(オシダ科)では背軸面からの発生が知られている[129]。また、アナコロプテリス Anachoropteris(†アナコロプテリス科[130])、ボトリオプテリス Botryopteris(†ボトリオプテリス科[130])、ホラゴケ属 Trichomanes(コケシノブ科)などでは葉柄の途中から、イヌチャセンシダ Asplenium tripteropus やトキワシダ Asplenium yoshinagae、オクタマシダ Asplenium pseudo-wilfordii(チャセンシダ科)、ヒメムカゴシダ Monachosorum arakii(コバノイシカグマ科)では羽片の基部中軸向軸側の付近からの不定芽が知られている[129]。



生理機能
光合成
葉は植物の体において、主な光合成の場となる[4]。光合成の代謝過程は葉の柵状組織と海綿状組織の葉肉細胞で起こる[131]。光合成を行う細胞の二酸化炭素の需要と、孔辺細胞による二酸化炭素の供給の協調作用が純CO2吸収として測定される光合成速度に影響する[131]。
光合成は葉の構造的特性と機能的特性に影響される[132]。葉の内部構造や葉の方向は光合成のための光吸収を最大化するようになっている[132]。また、葉は生育環境に対し馴化する[132]。
陸上植物は生育する環境の光条件に応じて形態的、生理的に異なった性質を持つ葉を作ることが多い[133]。弱光下で形成された葉を陰葉(いんよう、shade leaf)、強光下で形成された葉を陽葉(ようよう、sun leaf)という[133]。種によって陰葉と陽葉の分化の程度は異なる[133]。陰葉と陽葉のどちらが分化するかは、葉が発生するシュート頂ではなく既に成熟している葉に対する光環境で決まる[133]。
陰葉と陽葉には以下のような違いがある。
| 陰葉 | 陽葉 | |
|---|---|---|
| 形態的特徴 | (相対的に)面積が大きい[133] | (相対的に)面積が小さい[133] |
| 厚さが薄い[133][134][135] | 厚さが分厚い[133][134][135] | |
| 柵状組織の発達が悪い[133][134][99] | 柵状組織が発達し、多層になる[133][134][99] | |
| 生化学的特徴 | ||
| クロロフィルa/b比が小さい[133][134] | クロロフィルa/b比が大きい[133][134] | |
| 弱光下でも光合成効率をあげられるように、 反応中心あたりのクロロフィル量が多い[134] |
ルビスコを多く持ち、炭酸同化を増加させ、キサントフィルサイクルの構成要素の プールを大きくすることにより過剰エネルギーを放散する[134] | |
| 生理的特徴 | 光飽和時の葉面積当たりの光合成速度が小さい[133][135] | 光飽和時の葉面積当たりの光合成速度が大きい[133][135] |
| 葉面積当たりの呼吸速度が小さく、光補償点が低い[133] | 葉面積当たりの呼吸速度が大きく、光補償点が高い[133] |
単位葉面積当たりの重さを比葉重(ひようじゅう、LMA, leaf matter per area)といい、単位はg/m2である[135]。比葉重の大きな葉は物理的な強度が高い傾向にある[135]。陰葉より、強風などのストレスを受ける開けた環境の陽葉の方が比葉重が大きい[135]。また、草本植物に比べ木本植物の方が比葉重は大きく、木本の中でも落葉樹より常緑樹の方が比葉重が大きい[135]。常緑樹の葉は長い場合10年もの寿命を持つことがあり、長期間にわたって生存できるため、比葉重が大きい葉を作る[135]。
また、ギャップ形成などにより植物が置かれた環境が変わると、植物はその環境に適応する。葉が生育環境に適した性質を持つように生化学的および形態学的に調節された発生学的過程を馴化(順化、じゅんか、acclimation)という[134]。馴化は新たに展開する葉においても、既に成熟した葉においても起こりうる[134]。
落葉


葉は二次肥大成長を行わず、一定の季節に茎との境界に離層を分化して母体から脱落し、茎の表面に葉痕を残すことが多い[2]。温帯では、冬前に行うものが多いが、これを落葉という。落葉の有無により、落葉樹と常緑樹に区別する。落葉に際しては葉が枯れるので、黄色から茶色になることが一般的だが、特にはっきりとした色を発色するものがあり、黄色くなるものを黄葉、赤くなるのを紅葉という。
葉を落とす時期は、温帯では冬期の前が多いが、熱帯の乾燥地では、乾季の前に葉を落とす。また、常緑樹であっても、葉の寿命がくれば葉を落とす。葉の寿命は往々にして複数年にわたるが、温帯では、新芽が出る時期は初夏であり、この頃に古い葉を落とす例が多い。特殊な例としては、南西諸島等で植栽に用いられるのデイゴ(マメ科)は、花を咲かせる枝に限って葉を落とす。また、アコウ(クワ科)は、不定期に木全体の葉を落とし、新芽を出す。また、一般の落葉樹でも、落葉の時期でなくとも、乾燥がひどかったり、葉が塩害にあった時など、不特定の時期にも葉を落とす場合がある。
落葉を行う場合、葉柄のつけ根で葉がきれいに落ちるが、これは、ここに離層と呼ばれる切り離し面ができるためである。これは植物が自ら作るものである。押し葉標本を作ると葉がボロボロ落ちる木があるが、これも標本の枝中の水分が乾き切って死ぬ前に、離層を作ってしまうためである。したがって、葉を落とさないためには、枝を切り落としてすぐ、熱湯などで枝を殺してしまうとよいとも言う。
常緑植物でも一部の種、針葉樹のスギやニオイヒバ、メギ科のナンテン、ベンケイソウ科の多肉植物などで、冬には紅葉するが枯れて落葉はせず、春には再び緑色に戻るものがある。赤い色素は紫外線を吸収する作用があり、光合成活動が低下している時期に過剰な光による組織への悪影響(光阻害)を防止する効果があると考えられている。
用途
脚注
注釈
- 以降注記なしの立体ラテン文字は英名を示す。
- 複葉の各裂片も小葉 leaflet と呼ばれるが、別の用語である[1]。
- 茎から葉に繋がる維管束
- 環境条件によって異なる形態の葉を形成することをヘテロフィリー heterophylly、環境条件が一定でも成長過程で異なる形態の葉を形成することをヘテロブラスティー heteroblasty と呼び分け、それらを総称して「異形葉性」と呼ぶこともある[38]。
- 下記の鱗片葉とは異なる[69]
- 旧シロモジ属 Parabenzoin を含む
- ホメオボックス転写因子をコードする[109]。クラスⅠ KNOX 遺伝子とも[104]。
- syn. Aspidium cicutarium
出典
- 清水 2001, p. 119.
- 巌佐ほか 2013, p. 1069a.
- 長谷部 2020, p. 31.
- 清水 2001, p. 120.
- 清水 2001, p. 167.
- 長谷部 2020, p. 152.
- 西田 2017, p. 96.
- 西田 2017, p. 85.
- 長谷部 2020, p. 171.
- 日本植物学会 1990, p. 519.
- 伊藤 2012, p. 114.
- 長谷部 2020, p. 70.
- 西田 2017, pp. 93–94.
- 長谷部 2020, p. 71.
- 西田 2017, p. 92.
- 岩槻 1975, pp. 170–173.
- 長谷部 2020, pp. 153–157.
- Kenrick & Crane 1997, pp. 294–297.
- 長谷部 2020, pp. 158–159.
- 西田 2017, p. 155.
- 加藤 1999, pp. 28–29.
- 西田 2017, pp. 148–154.
- 西田 2017, p. 91.
- 長谷部 2020, p. 144.
- 西田 2017, p. 88.
- 西田 2017, p. 158.
- 長谷部 2020, p. 146.
- 長谷部 2020, p. 148.
- Taiz & Zeiger 2017, p. 553.
- Taiz & Zeiger 2017, p. 558.
- 長谷部 2020, p. 176.
- 西田 2017, p. 156.
- 長谷部 2020, p. 128.
- Kenrick & Crane 1997, pp. 288–292.
- ギフォード & フォスター 2002, p. 101.
- 原 1994, p. 36.
- 清水 2001, p. 164.
- 長谷部 2020, p. 184.
- 巌佐ほか 2013, p. 62f.
- 清水 2001, p. 122.
- 原 1994, p. 37.
- 清水 2001, p. 142.
- 清水 2001, pp. 23–24.
- 原 1994, p. 42.
- 岩瀬 & 大野 2004, p. 74.
- 巌佐ほか 2013, p. 896i.
- 巌佐ほか 2013, p. 1200h.
- 清水 2001, p. 126.
- 清水 2001, p. 132.
- 巌佐ほか 2013, p. 329c.
- 巌佐ほか 2013, p. 1424e.
- 清水 2001, p. 276.
- 清水 2001, p. 127.
- 清水 2001, p. 124.
- 清水 2001, p. 152.
- 清水 2001, p. 140.
- 熊沢 1979, p. 157.
- 小倉 1954, p. 139.
- 熊沢 1979, p. 158.
- 熊沢 1979, p. 159.
- ギフォード & フォスター 2002, p. 405.
- 長谷部 2020, p. 199.
- Christopher J. Earle. “Gymnosperms”. The Gymnosperm Database. 2023年6月29日閲覧。
- ギフォード & フォスター 2002, p. 413.
- ギフォード & フォスター 2002, p. 414.
- 長谷部 2020, p. 209.
- 岩瀬 & 大野 2004, p. 75.
- 岩瀬 & 大野 2004, p. 76.
- 清水 2001, p. 144.
- 清水 2001, p. 148.
- 清水 2001, p. 26.
- 原 1994, p. 44.
- 小倉 1954, p. 144.
- 加藤 1997, p. 80.
- 清水 2001, p. 108.
- 熊沢 1979, pp. 31–32.
- ギフォード & フォスター 2002, p. 493.
- 熊沢 1979, p. 262.
- 原 1994, p. 67.
- 原 1994, p. 43.
- 清水 2001, p. 146.
- 熊沢 1979, p. 232.
- 小倉 1954, p. 145.
- 巌佐ほか 2013, p. 929e.
- 熊沢 1979, p. 261.
- 長谷部 2020, p. 59.
- 清水 2001, p. 130.
- 清水 2001, p. 202.
- 巌佐ほか 2013, p. 1425a.
- 原 1994, p. 59.
- 清水 2001, p. 158.
- 原 1994, p. 95.
- 原 1972, p. 178.
- 原 1972, p. 179.
- 長谷部 2020, p. 89.
- 巌佐ほか 2013, p. 1426d.
- 原 1994, p. 61.
- 清水 2001, p. 160.
- 原 1972, p. 180.
- 原 1994, p. 65.
- 清水 2001, p. 161.
- 原 1994, p. 66.
- 巌佐ほか 2013, p. 683e.
- 巌佐ほか 2013, p. 1422b.
- Taiz & Zeiger 2017, p. 555.
- 原 1972, p. 72.
- 原 1972, p. 73.
- 原 1994, p. 143.
- 長谷部 2020, p. 145.
- 長谷部 2020, p. 85.
- 長谷部 2020, p. 162.
- ギフォード & フォスター 2002, pp. 253–254.
- 岩槻 1992, p. 12.
- 長谷部 2020, p. 34.
- 巌佐ほか 2013, p. 653b.
- 清水 2001, p. 213.
- 清水 2001, p. 216.
- 巌佐ほか 2013, p. 1423f.
- 熊沢 1979, p. 181.
- 清水 2001, p. 224.
- 熊沢 1979, p. 172.
- 巌佐ほか 2013, p. 1207e.
- 清水 2001, p. 222.
- 熊沢 1979, p. 177.
- 巌佐ほか 2013, p. 1207h.
- 熊沢 1979, p. 178.
- 熊沢 1979, p. 174.
- 熊沢 1979, p. 175.
- 熊沢 1979, p. 176.
- 西田 2017, p. 295.
- Taiz & Zeiger 2017, p. 245.
- Taiz & Zeiger 2017, p. 246.
- 巌佐ほか 2013, p. 100d.
- Taiz & Zeiger 2017, p. 249.
- 塩井, 井上 & 近藤 2009, p. 270.
参考文献
- Kenrick, Paul; Crane, Peter R. (1997). The Origin and Early Diversification of Land Plants —A Cladistic Study. Smithonian Institution Press. ISBN 1-56098-729-4
- Laubenfels (1953). “The external morphology of coniferous leaves”. Phytomorphology 3: 1–20.
- 伊藤元己『植物の系統と進化』裳華房〈新・生命科学シリーズ〉、2012年5月25日。ISBN 978-4785358525。
- 岩槻邦男『日本の野生植物 シダ』平凡社、1992年2月4日。ISBN 4-582-53506-2。
- 巌佐庸、倉谷滋、斎藤成也、塚谷裕一『岩波生物学辞典 第5版』岩波書店、2013年2月26日。ISBN 9784000803144。
- 岩瀬徹、大野啓一『野外観察ハンドブック 写真で見る植物用語』全国農村教育協会、2004年5月3日。ISBN 4-88137-107-X。
- 小倉謙『植物解剖及形態学』(第4版)養賢堂、1954年4月15日。
- 岩槻邦男 著「シダ植物門」、山岸高旺 編『植物分類の基礎』(2版)図鑑の北隆館、1975年5月15日、157-193頁。
- 加藤雅啓『植物の多様性と系統』1997年。
- 加藤雅啓『植物の進化形態学』東京大学出版会、1999年5月20日、60-82頁。
- アーネスト M. ギフォード、エイドリアンス S. フォスター『維管束植物の形態と進化 原著第3版』長谷部光泰、鈴木武、植田邦彦監訳、文一総合出版、2002年4月10日。ISBN 4-8299-2160-9。
- 熊沢正夫『植物器官学』裳華房、1979年8月20日。ISBN 978-4785358068。
- 塩井祐三、井上弘、近藤矩朗『ベーシックマスター 植物生理学』オーム社、2009年2月20日。ISBN 978-4274206634。
- 清水建美『図説 植物用語事典』八坂書房、2001年7月30日、132頁。ISBN 4-89694-479-8。
- リンカーン・テイツ (Lincoln Taiz)、エドゥアルト・ザイガー (Eduardo Zeiger)、イアン・M・モーラー (Ian Max Møller)、アンガス・マーフィー (Angus Murphy) 著、西谷和彦、島崎研一郎 訳『テイツ/ザイガー 植物生理学・発生学 原著第6版 (原著:Plant Physiology and Development, Sixth Edition)』講談社、2017年2月24日(原著2015年)。ISBN 978-4-06-153896-2。
- 原襄『植物の形態』裳華房〈基礎生物学選書〉、1972年11月25日。
- 原襄『植物形態学』朝倉書店、1994年7月16日。ISBN 978-4254170863。
- 西田治文『化石の植物学 ―時空を旅する自然史』東京大学出版会、2017年6月24日。ISBN 978-4130602518。
- 日本植物学会『学術用語集 植物学編(増訂版)』文部省、1990年3月20日。ISBN 462103376X。
- 長谷部光泰『陸上植物の形態と進化』裳華房、2020年7月1日。ISBN 978-4785358716。
