流用 (生物学)
生物学における流用[1](りゅうよう、co-option、コ・オプション[2][3])とは、発生遺伝子系の進化や進化的に新しいパターンの獲得において、既存の遺伝子系(遺伝子群やそれに基づく形質・発生プログラムなど)が別の場所や別の時間で用いられることでそれまで祖先に存在しなかった構造をもたらすことである[1][2][4]。進化発生学の場面では、特に特定のツールキット遺伝子群の発現ドメインが祖先には存在しなかった場所にもたらされることを指してコ・オプションという[2]。
対して、祖先が持たない新しい遺伝子系を進化させることを「新生 de novo」と呼ぶ[1]。流用と新生には中間的なものもあり、遺伝子系に含まれる遺伝子やそれらの制御関係が一部のみ変わっている場合も存在する[1]。
概要
古典的形態学において、あらゆる構造物は新しく現れないとされるが、イルカや魚竜の背鰭や、古生代の魚類の胸鰭のように、例外的に突然誕生した構造物を「新規形質」や「新形成物」と呼び特別扱いしてきた[5]。流用により、このような構造の出現の原理を説明することができる[5]。一連の遺伝子制御系である発生過程の中には階層性があって、多くの遺伝子群からなる一つのセットのようないくつかの発生上の機能的単位(まとまり)が認められることがあるが、この機能的単位をモジュールと呼ぶ[1][5]。流用では、このモジュールがそのまま新しい場所に移植される[5]。モジュールはしばしば遺伝子制御のネットワークやタンパク質間の相互作用をベースに組みあがっている発生プログラムのサブルーチンと呼べるものであり、進化を通じて保存される分子レベルでの相同性を基盤として成立している[5]。つまり、ある遺伝子が別に遺伝子を発現させるという関係が進化的に保存され、そのつながりが発現ネットワークを形成している[5]。そのため、遺伝子発現ネットワークの中で上流にある遺伝子の発現部位が新しく追加されれば、その下流にある同じモジュールの因子群が発現され、そこに新しいパターンをもたらすことで新たな器官が獲得される[5]。このようなメカニズムであるため、比較的短期間の進化も可能であり、形態構造では新規であるが発生の分子的機構では相同性を見出すことができる[5]。しかし、分子発生的プログラムは相同であっても、器官自体は全く別のイベントとして進化してきたものであり、形態的なボディプランの相同とは異なる[5]。
その構造がどこに存在するか、ということを位置価と呼ぶ[6]。ある器官の発生において、その一部の構造を構成する細胞群は発生のある段階で自分の発生運命を他の構造とは別のものとして知らねばならず、それは器官原基の正しい相対的位置に付随していなければならない[6]。この位置決定システムをある軸において指定するのが Hox遺伝子群の発現パターンである[6]。
全く新しい形質や遺伝子プログラムではなく、祖先において存在していたものの転用という意味で、外適応(イグザプテーション、exaptation)を流用の類似概念とする場合があるが、厳密に分けるべきだとの考えもある[2]。外適応は、それ自体が新しいわけではないという意味でむしろ鍵革新(key innovation)に近く、流用は比較発生学や比較形態学の方法では説明できないタイプの進化的形態変化の説明に限定するべきだとされる[2]。また、流用はもっぱら派生的な形質の成立を説明するもので、互いに離れた分類群の基本的なボディプランの構成要素に見る類似性が真に祖先的形質の共有なのか、あるいは何らかの祖先的形質が異なった機能や形質の獲得のために流用されたものなのか、直ちに判断できない事例は多い[2]。
植物における流用
胞子体世代
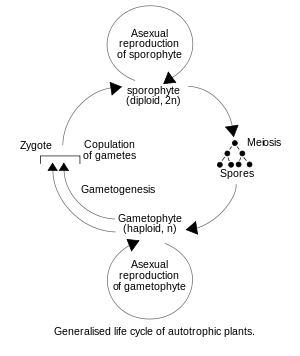
陸上植物は単相単世代型の隔膜形成体緑藻類[注釈 1]とは違い、単複相世代交代型を進化させてきたため、その生活環には単相の配偶体だけでなく、複相の胞子体世代を持つようになった[8]。この胞子体世代の発生を司る遺伝子系として、流用仮説と新生仮説の2つの仮説が提唱されていた[9]。前者は配偶体世代の発生の遺伝子系の流用により胞子体世代の発生の仕組みが進化したとする仮説で、後者は胞子体世代で全く新しい発生の仕組みを獲得したとする仮説である[9]。現在は研究が進み、実際にはどちらの仕組みも用いられていることが分かった[9]。
例えば、新生の例としては次のようなものがある。被子植物のシロイヌナズナの LFY (LEAFY) 転写因子は、花器官形成遺伝子である MADS-box 遺伝子の転写を抑制し、花器官[注釈 2]を形成する[9]。対してコケ植物のヒメツリガネゴケは LFY 遺伝子と MADS-box 遺伝子のオルソログ遺伝子を持っているものの、 LFY は MADS-box 遺伝子を転写制御していない[9]。そのため、 LFY が MADS-box 遺伝子を制御する遺伝子系はコケ植物が被子植物と分岐した後に、被子植物の系統で新生したことになる[9]。
対して、胞子体のもつ根毛は配偶体の仮根の形成に用いられる遺伝子系の流用だと考えられている。被子植物の胞子体の根毛形成に用いられているのと似た遺伝子系はヒメツリガネゴケの配偶体の仮根形成に用いられている[9]。そのため、被子植物と蘚類の共通祖先で用いられていたメカニズムが、ヒメツリガネゴケでは仮根形成に、被子植物では根毛形成に流用された可能性が高い[9]。
リグニン生合成
コケ植物ではリグニンを持たないのに対し、維管束植物においてリグニンが二次細胞壁に蓄積し植物体の強度を高め大型化する[10]。しかし、コケ植物のヒメツリガネゴケにおいてもリグニン生合成系の遺伝子は最終産物であるリグナンを重合させる酵素以外は全て持っており、この前駆的リグニン生合成系はクチクラの疎水性細胞外生体高分子の生合成に寄与している[10]。また、この高分子は隔膜形成体植物(フラグモプラスト植物)の共通祖先で進化したスポロポレニン生合成系から進化してきたと考えられている[10]。これらのことから、隔膜形成体緑藻類では休眠性受精卵を保護しているスポロポレニン生合成系が陸上植物の共通祖先において単複相世代交代が進化した際に胞子で用いられ、それがクチクラ生合成の遺伝子系の進化により乾燥した環境(陸上)での生活を可能にし、その後維管束植物の共通祖先でスポロポレニン生合成系やクチクラ生合成系を流用することでリグニン生合成系を獲得したと考えられている[10]。
モノフィレアの花序
_(15481094276).jpg.webp)
イワタバコ科のモノフィレア属 Monophyllaeaは、葉の上に分裂組織ができ、花序を形成する[11]。これは茎に花をつける普通の体制を持つイワタバコ科の植物における花序分裂組織形成の発生遺伝子系を葉の上に異所的流用したものだと考えられる[11]。
複葉形成
複葉形成には、茎頂分裂組織で使われているものと似た遺伝子系が用いられている[1]。これは KNOX-CUC とよばれるモジュールが発生過程で流用され、繰り返し用いられていると考えられている[1]。
内珠皮形成
被子植物の胚珠を取り囲む珠皮は内珠皮と外珠皮の二層からなる[12]。シロイヌナズナの胚珠では、茎頂分裂組織の肝細胞維持に機能する WUS (WUSCHEL)が発現しており、WUS がないと珠皮ができず、WUS を異所的に発現させると異所的に珠皮ができるため、WUS が茎頂分裂組織様の性質を胚珠原基に与え、茎頂分裂組織から側生器官として葉ができるのと同様に珠皮が形成されるのではないかと推定されている[13]。内珠皮も外珠皮も葉の向軸側と背軸側を決定する遺伝子やそのパラログが発現している[13]。これらのことから、茎頂分裂組織で葉を形成する仕組みを雌性胞子嚢が形成されていた位置で発現させることで、珠皮が進化した可能性がある[13]。
動物における流用
顎口類の対鰭
脊椎動物を含む左右相称動物の前後軸を特異化する Hoxコードの一部が顎口類の肢芽および鰭芽の極性化に流用されている[2][6]。
対鰭およびそこから派生した四肢は顎口類の一部にのみ獲得された派生形質であり、対鰭の原基である鰭芽および四肢の原基である肢芽の前後軸と遠近軸に沿って、細胞群の位置価を決定する遺伝子群(HoxA クラスターと HoxD クラスター の遺伝子)がいくつか入れ子状となる発現パターンを示す[6]。遠位から近位を通して Hoxa9 が発現し、中位から近位にはそれに加えて Hoxa10 と Hoxa11が、さらに最も遠位ではそれらに加えて Hoxa12 および Hoxa13 が発現する[6]。また、前後軸方向に、領域の多くを Hoxd9 が、より後方ではそれに加えて Hoxd10 と Hoxd11が、さらに最も後方ではそれらに加えて Hoxd12 および Hoxd13 が発現する[6]。同様のものが左右相称動物の前後軸において発現し、胚の各部の位置価を決定する Hoxコードであり、手足とは違い広範に、時期も早く発現する重要な発生プログラムである[6]。そのため、体軸の前後を規定する発生機構が流用され、対鰭の発生プログラムとなったと考えられている[6]。
また化石記録から胸鰭は腹鰭より起源が古いと考えられている[14]。例えば、オストラコダームでは胸鰭は持つが腹鰭は持たない[14]。そのため、胸鰭と腹鰭およびそれに由来する前肢と後肢はそれぞれ単純な系列相同物ではなく、胸鰭の進化で獲得した対鰭の発生モジュールを流用し、進化のある時点で後方に二次的に付加して腹鰭となり、それを進化させて四肢動物では四肢として用いているということが推定される[14]。
カメの甲羅
カメの甲羅は上記の肢芽のパターン形成に寄与する Wnt5a シグナリングシステムと類似したものによりパターン形成される[14]。甲羅を持たない他の脊椎動物胚ではこのシグナル機構が背で働くことはないため、カメの甲羅という新規形態の獲得はこの流用であると考えられている[14]。
甲虫の角の形成

甲虫類の雄で発達する角は他の昆虫では角原基に相同な器官が存在せず、何もないところからいきなり新しい構造が得られるように見えるが、これも流用による[3]。角のパターン形成において、角の構造の根元から先端にかけて1セットの遺伝子が位置特異的に発現する[3]。その中で中心的機能を持つのは遠位に発現する節足動物で普遍的に付属肢のパターン形成に関与する Distalless (Dll) 遺伝子であり、角のパターン形成は付属肢のパターン形成に用いられる遺伝子セットの流用である[3][2]。付属肢では、最も近位で hth (homothorax), n-exd (extradenticle) が、最も遠位および hth, n-exd の働く部分のうち遠位部分で Dll が、その間で両者とやや重複して dac (dachshund) が発現してパターンが形成されるが、昆虫の角では近位から遠位まで通して dac が、最も近位のみで hth, n-exd が、中位から最遠位にかけて al が、そして最も遠位で Dll が発現することで角のパターンが形成されている[3][15]。
脚注
出典
- 長谷部 2020, p. 148.
- 巌佐ほか 2013, p. 469d「コ・オプション」
- 倉谷 2015, pp. 123–124.
- 倉谷 2015, p. 123.
- 倉谷 2015, pp. 124–126.
- 倉谷 2015, pp. 127–128.
- 長谷部 2020, p. 8.
- 長谷部 2020, pp. 25–26.
- 長谷部 2020, pp. 78–79.
- 長谷部 2020, pp. 112–113.
- 長谷部 2020, pp. 168–169.
- 巌佐ほか 2013, p. 1080b「胚珠」
- 長谷部 2020, pp. 180–190.
- 倉谷 2015, pp. 128–129.
- Moczek, Armin P.; Rose, Debra; Sewell, William; Kesselring, Bethany R. (2006). “Conservation, innovation, and the evolution of horned beetle diversity”. Development Genes and Evolution 216 (11): 655-665. doi:10.1007/s00427-006-0087-2.