視覚野
視覚野(しかくや、英: visual cortex)という用語は、V1と略される一次視覚野 (または、線条皮質 (striate cortex、有線皮質とも)) 及びV2、V3、V4、V5と略される外線条皮質 (extrastriate cortex、有線外皮質、有線領外皮質とも) を示す。一次視覚野はブロードマンの脳地図における17野と解剖学的に同等である。
| 脳: 視覚野 | |
|---|---|
 ブロードマンの脳地図における17野 (一次視覚野) が赤、18野がオレンジ、19野が黄色で示されている。 | |
| 名称 | |
| 日本語 | 視覚野 |
| 英語 | Visual area, Visual cortex |
| 関連構造 | |
| 上位構造 | 大脳皮質、後頭葉 |
| 関連情報 | |
| Brede Database | 階層関係、座標情報 |
| MeSH | Visual+Cortex |
イントロダクション
一次視覚野 (V1) は、後頭葉にある鳥距溝の周りに位置する顆粒皮質 (Koniocortex) であり、外側膝状核から直接情報を受け取る。

V1 は背側皮質視覚路と腹側皮質視覚路と呼ばれる2種類の主要な視覚経路に情報を伝える。
- 背側皮質視覚路は V1より始まり V2、背内側野、MT 野 (V5 とも) を通過し、後頭頂皮質へと向かう。背側皮質視覚路は時に、where経路とも呼ばれ、運動、物体の位置や、眼や腕の制御、特に視覚情報を用いたサッカードや到達運動に関連付けられている[1]。
- 腹側皮質視覚路は V1より始まり V2、V4、を通過し下側頭皮質へと向かう。腹側皮質視覚経路は時に、what経路とも呼ばれ視覚対象の認識や形状の表象(意識にのぼる映像)と関連している。また、長期記憶の貯蔵とも関連している。
背側皮質視覚路と腹側皮質視覚路の二分法 (またはwhere/whatの二分法、行動 (action) /知覚 (perception) の二分法) [1]はアンガーライダー (Ungerleider) とミシュキン (Mishkin) によって最初に定義され[2]、現在でも視覚神経科学者や心理学者の間で論争を巻き起こしている。おそらく、この二分法は視覚野での実際の出来事を単純化し過ぎている。この研究はエビングハウス錯視などによって知覚の判断が歪められるという発見に基づいているが、通常、人が把握運動などの行動によって反応する時、知覚の歪みは発生しない。しかし、最近の研究 [3]では行動と知覚はこれらの錯視で同様に錯覚が起きているとされている。
視覚野のニューロンは、視覚刺激が受容野に呈示された時、活動電位を発生させる。定義から、受容野は全視野の内、活動電位を生じさせるものを指す。しかし、あるニューロンに関しては、そのニューロンの受容野の特定の刺激のサブセットに応答する。この特性はチューニングと呼ばれる。より初期の視覚野ではより単純なチューニングが行われている。例えば、V1 のあるニューロンはその受容野に現れる垂直な線に対して発火する。より高次の視覚野ではより複雑なチューニングが行われている。例えば下側頭皮質 (IT) のあるニューロンはその受容野に現れる特定の顔に対して発火する。
視覚野は後大脳動脈の鳥距溝枝から血液の供給を最も受ける。
最近の研究
一次視覚野の研究では活動電位をネコ、フェレット、ラット、ネズミ、サルの脳に刺した電極によって計測したり、動物における内因性光学信号を計測したものや、ヒトやサルの V1 のfMRI信号を計測するものがある。
最近のある発見では、ヒトの V1 のfMRI により計測される信号は注意による調節 (attentional modulation) を強くうけるというものがある。この結果はマカクザルの生理学的な研究において、注意による調節によりニューロンの発火に殆ど変化が見られなかった結果と対照的である。しかし、マカクザルの研究は1ニューロンのスパイク活性を調べたものであったが、fMRI 信号の神経基盤は主に後シナプス増強 (PSP) によるものである。したがって、今回の結果の違いがそのままヒトとマカクの生理学的な違いを意味するものではない。
他の最近の V1 の研究では、そのチューニングの特性を完全に説明し、皮質のカノニカル回路のモデルとして利用しようと試みたものがある。
一次視覚野の損傷は暗点 (視野にできた『穴』) を生み出す。興味深いことに、暗点を持つ患者は意識的にそれを知覚出来ないにも関わらず、暗点における視覚情報を利用することが出来る。この現象は盲視と呼ばれ、意識に相関した脳活動 (neural correlate of consciousness) に興味を持つ研究者に広く研究されている。
一次視覚野 (V1)

一次視覚野は脳の視覚野で最も良く調べられている領域である。研究されたすべての哺乳類において、一次視覚野は後頭葉 (後頭葉は視覚刺激の処理を担っている) の後頭極に位置している。一次視覚野は、最も単純で最も初期に活動する視覚野で、静止、または運動する対象に関する情報の処理に特化し、また、パターン認識に力を発揮する。
機能的に定義された一次視覚野と呼ばれる領域は、解剖学的に定義される線条皮質とおよそ同等である。"線条皮質 (striate cortex)"という名前はジェンナリ線 (stria of Gennari) と呼ばれる、外側膝状体から灰白質のIV層へと伸びる髄鞘化された軸索の、肉眼でも識別可能な大きな縞模様に由来する。
一次視覚野はI層からVI層までラベルされた6層の機能的に異なる層に分けられる。その内、外側膝状体 (LGN) から殆どの視覚情報の入力を受けるIV層は4A、4B、4Cα、4Cβ のさらに4層に分けられる。4Cα 亜層は外側膝状体からの殆どの巨細胞性入力を受け、4Cβ 層は小細胞性経路からの入力を受ける。
ヒト成人の一次視覚野の平均ニューロン数はそれぞれの大脳半球につき、約1億4000万個ほどであると見積もられる。(Leuba & Kraftsik, Anatomy and Embryology, 1994)
機能

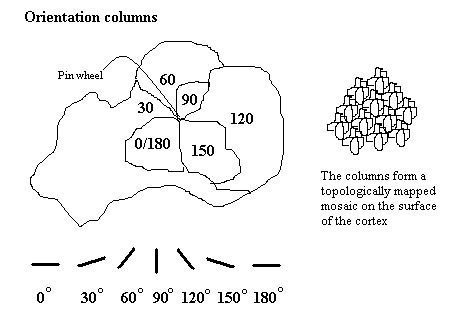
V1 は視覚における空間情報の明確なマップを持つ。例えば、ヒトの鳥距溝上壁は視野の下半分の視覚情報に強く応答し、下壁は視野の上半分に応答する。概念的には、このレチノトピーのマッピングは網膜から V1 へ視覚像の変換とみなせる。視野と V1 における位置の対応は非常に正確で、盲点でさえも V1 でマップされている。この対応関係は進化的に基本的で、V1 をもつ殆どの動物において見られる。中心窩を持つ動物やヒトでは、V1 の大きな割合を占める領域が、視野の中心にある小さい領域をマップしている。この現象は皮質拡大(cortical magnification) として知られる。おそらく、正確な空間的コーディングのために、V1 のニューロンの受容野の大きさは全ての視覚野の微小領域で最小であろう。
V1 ニューロンのチューニング特性 (ニューロンの応答選択性) は時間によって大きく異なる。短時間 (40ms前後) で、それぞれの V1 ニューロンは刺激の小さな組に対して強くチューニングされる。つまり、それぞれのニューロンの応答性は視覚刺激の方位、空間周波数や色の僅かな違いに分けることが出来る。さらに、ヒトと、両眼視をする動物の、それぞれの V1 ニューロンは眼優位性を持ち、文字通り両眼のうちの片方に対しチューニングしている。V1 と一般的な一次感覚野のニューロンは似たチューニング特性を持つもので集まり、皮質コラムと呼ばれる構造を作る。デイヴィッド・ヒューベルとトルステン・ウィーセルは眼優位性と方位の2つのチューニング特性の皮質コラムについて古典的なモデル (the classic ice-cube organization model) を提唱した。しかし、このモデルは色や空間周波数などの、ニューロンがチューニングされる他の多くの特徴については適合しなかった。これら全ての V1 の皮質コラムの正確な構成は、最近の研究において、いまだにホットなトピックである。
現在では、V1 ニューロンの初期の応答は選択的な時空間フィルター (spatiotemporal filters) で構成されていると考えられている。空間次元では、V1 の機能は多くの空間的に局所的で複合的なフーリエ変換に似たものであると考えられる。理論的には、このようなフィルターは、空間周波数や方位、運動、運動方向、速度 (したがって、時間周波数も) やほかの多くの時空間的特徴について神経的な処理を行うことができる。V1 ニューロンにおける実験は、この理論を実証するが、さらに新しい疑問も生む。
時間的に遅く (100 ms後) に、V1 のニューロンはシーン (scene) のより大局的な構造に応答する(Lamme & Roelfsema, 2000)。このような応答特性は回帰性の処理 (高次の領域から低次の領域への影響) と錐体細胞の水平な結合 (Hupe et al 1998) により生じるものと考えられる。
V1 へと中継される視覚情報は空間的 (または光学的)な像としてというよりは、局所的なコントラストとしてコードされている。例えば、片側が黒、反対側が白から成る像は、局所的なコントラストが最大である黒と白を分ける線がコードされ、輝度の情報 (黒さや白さ自体) は僅かなニューロンしかコードしない。情報が高次の視覚領域へと中継されるにしたがって、非局所的な周波数/位相信号がより多くコードされていく。重要なことは、このような皮質での視覚処理の初期では、視覚情報の空間的な位置は局所的なコントラストのコーディングのなかで強く保存されている点である。
V2
V2は前有線皮質 (prestriate cortex) とも呼ばれ[4]、視覚野の2番目に広い領域で、視覚連合野の1番初めの領域である。この領域は V1 からの強いフィードフォワード接続を受け、V3、V4、V5へと接続を送っている。また、V1 への強いフィードバック接続を持つ。
解剖学的には V2 は左右半球の背側と腹側の4つの四分円に分けられる。この4領域が一緒になって視覚世界の完全なマップを形作っている。機能的には V2 は V1 と多くの共通な特性を持つ。細胞は方向や空間周波数、色などの単純な特性にチューニングされている。V2 ニューロンの多くの応答は主観的輪郭の向きやその視覚刺激が図の一部か地の一部かなどのより複雑な特性によって調節を受けている。(Qiu and von der Heydt, 2005).
最近の研究によって、V2 の細胞は注意による調節 (attentional modulation) が小さく (V1 より大きく、V4 より小さい)、また、単一の受容野における異なる下位領域の、複数の方向に反応することで、適度に複雑なパターンにチューニングされていることが判明している。
V3 を含む3次視覚皮質複合体
3次視覚皮質複合体という用語は V2 の前部に接する皮質領域を示し、V3領域を含んでいる。ヒトにおいて、"複合体"という名前は V3 と呼ばれる領域の正確な範囲について、V2 の前にある領域は2、3の機能的な下位領域を含んでいるとする研究者によって論争が起きているという事実による。例えばデイヴィット・ヴァン・エッセン (David Van Essen) ら (1986) は大脳半球の上部に "背側 V3" (dorsal V3) という領域が存在し、脳の下部に位置する"腹側 V3" (ventral V3, 腹側後部領域, ventral posterior area, VP) とは異なると提唱している。背側 V3 と腹側 V3 は脳の異なる領域から異なる接続を受け、多くの染色法によってことなる染まり方をし、視覚刺激の異なる組み合わせ (例えば色選択的ニューロンは腹側 V3 に多い) に応答する。他の下位領域として、V3A と V3B がヒトにおいて報告されている。これらの下位領域は、背側 V3 の付近に存在するが V2 とは区別される。
背側 V3 は通常、背側皮質視覚路の一部と考えられ、V2 と一次視覚野から入力を受け、後頭頂葉に投射している。解剖学的にはブロードマンの脳地図における19野に位置する。fMRI による最近の研究によって V3/V3A 領域は広域的な動きの処理に関わるとされている[5]。背側 V3 を全視野を表現する背内側野 (DM) と呼ばれる大きな領域の一部と考える他の研究も存在する。背内側野のニューロンは視野の大部分を占める大きなパターンの同期した動きに応答する。(Lui and collaborators, 2006)
腹側 V3 (VP)は一次視覚野からの非常に弱い結合と、下側頭回との強い結合を持つ。初期の研究では、腹側 V3 は視野の上半分 (固視点の上側) を表現している領域のみを指していた。しかし、より最近の研究では、腹側 V3 は以前考えられていたよりもより広く、他の視覚野と同様に視野全体を表現する領域を考えられている。この改訂されたより広い腹側 V3 はローサ (Rosa) とツィーデル (Tweedale) によって腹外側後部領域 (VLP) と呼ばれた[6]。
V4
V4はマカクザルでは、有線外視覚皮質に存在する視覚野の一部である。V4 は V2 の前部に位置し、後下側頭野 (PIT) の後部に位置する。V4 は左右のV4d、V4vの4領域から成るとされ、さらに尾側と吻側の下位領域を加える者もいる。ヒトにおける V4 ホモログはまだ知られておらず、精査されている事柄である。
V4 は腹側皮質視覚路の3番目の領域で、V2 からの強いフィードフォワードな入力を受け, 後下側頭野 (PIT) へと強い結合を持つ。また V1 からの直接の入力をその中心部へ受けている。加えて、V5 と背側前月状回 (DP) へと弱い結合が存在する。
V4 は腹側皮質視覚路において強い注意による調節 (attentional modulation) を受ける最初の領域である。多くの研究では、選択的注意が V4 の発火頻度を20%変化させるとしている。モラン (Moran) とデシモーネ (Desimone) による、影響力の大きい論文は、このような効果を特徴付け、注意による効果は視覚野のどこにおいても見られることを明らかにした最初の論文である[7]。
V1 のように V4 は方向、空間周波数、色にチューンしているが、V1 とは異なり、V4 は対象の特徴の中間的な複合性、例えば、単純な幾何学的形状などに、チューンしている。しかし、V4 のチューニング空間の完全なパラメーターの記述を行った者はいない。また、V4 は下側頭回のように顔などの複雑な対象にはチューニングされていない。
V4 の発火特性は1970年代後半にこの領域の名付け親でもあるセミール・ゼキ (Semir Zeki) によって初めて描写された。それまでは、V4 はその解剖学的な名である前月状回として知られていた。元々、ゼキは V4 の目的は色情報を処理するためであると考えていたが、1980年代前半に V4 は他のより初期の視覚野と同様に形の認知に直接関わっていると示された。この研究はアンガーライダー (Ungerleider) とミシュキン (Mishkin) により1982年に初めて提唱された2経路仮説 (上述) によっても支持されている。
最近の研究によって、V4 は刺激の顕著性 (salience) をコードする長期可塑性を示し、その可塑性は前頭眼野からの信号によってコントロールされることが分かった。これにより、注意によって受容野の空間的な特性が変化する。
V5/MT野
V5はMT野 (middle temporal) としても知られ、外線条野の一部であり、運動の知覚、局所運動信号のグローバルな知覚への統合、及びある種の眼球運動における主要な役割を担うと考えられている[8]。
神経結合
MT 野は様々な皮質及び皮質下の脳領域に結合している。その入力は V1、V2、背側 V3、背内側野[9] [10]、外側膝状体の顆粒細胞 (koniocellular) 領域[11]、及び、下視床枕から受けている。MT 野への投射のパターンは視野の中心と周辺で幾分異なっている。後者では、皮質正中部と脳梁膨大後部からの入力を受ける[12]。
一般的には、MT 野への"最も重要な"入力はV1に由来すると考えられている[8]。しかし、いくつかの研究は MT 野のニューロンは V1 ニューロンが破壊または不活性化された後でも、視覚情報に対して、多くの場合は方向選択的に応答できることを示した[13]。加えて、セキール・ゼキ (Semir Zeki) らによる研究ではある種の視覚情報は V1 に到着する前に MT 野に到着することが示唆されている。
MT 野はその主な出力を、FST や MST、V4t (中側頭半月部) を含むその領域を直接取り囲む領域へと送っている。他の MT 野の出力先は眼球運動に関係する前頭葉と頭頂葉の領域 (前頭眼野と外側頭頂間野 (LIP)) である。
機能
MT 野のニューロンの電気生理学的な特性を調べた最初の研究により、大部分の細胞が運動する視覚刺激の速度と方向にチューニングされていることが分かった[14] [15]。このような結果は MT 野が視覚運動の処理に重要な役割を担っていることを示唆している。
MT 野の損傷による研究によっても、その運動知覚と眼球運動における役割は示されている。また、運動を知覚することが出来ず、世界が静的な"フレーム"の連続に感じられる患者の神経心理学的研究により、霊長類における MT 野のヒトにおけるホモログは V5 であると分かった[16][17]。
しかし、V1 のニューロンも運動の方向や速度にチューニングされていることから、これらの初期の結果は、V1 では処理不可能な何を MT 野では処理可能なのかという問題をまだ解けてはいない。多くの研究によって、この領域が局所的な視覚運動信号を複雑な対象の大局的運動 (global motion) へと統合していることが分かった[18]。 例えば V5 の損傷により、運動の知覚と複雑な刺激の処理に障害が起きる。V5 には複雑な視覚的特徴 (線の端やコーナーなど) の運動に選択的に応答する多くのニューロンがある。V5 にあるニューロンの微小刺激は動きの知覚に影響を及ぼす。例えば、上向きの運動に選択的に応答するニューロンを見つけて、電極により刺激したとき、ニューロンを刺激されたサルは上向きの運動をより多く報告しやすくなる[19]。
MT 野で行われている計算の正確な形に関してはいまだに論争が存在する[20]。そして、ある研究では、特徴の運動は V1 のような視覚野における低次の領域でも、実はすでに利用可能であるとしている[21] [22]。
参考文献
- Goodale & Milner (1992). “Separate pathways for perception and action.”. Trends in Neuroscience 15: 20-25. doi:10.1016/0166-2236(92)90344-8.
- Ungerleider and Mishkin (1982). Ingle DJ, Goodale MA and Mansfield RJW. ed. Analysis of Visual Behavior. MIT Press
- Franz VH, Scharnowski F, Gegenfurtner (2005). “Illusion effects on grasping are temporally constant not dynamic.”. J Exp Psychol Hum Percept Perform. 31(6): 1359-78.
- Gazzaniga, Ivry & Mangun: Cognitive neuroscience, 2002
- Braddick, OJ, O'Brian, JMD, et al (2001). “Brain areas sensitive to visual motion.”. Perception 30: 61-72. doi:10.1068/p3048.
- Rosa MG, Tweedale R (2000) Visual areas in lateral and ventral extrastriate cortices of the marmoset monkey. J Comp Neurol 422:621-51.
- Moran & Desimone. Selective Attention Gates Visual Processing in the Extrastriate Cortex. Science 229(4715), 1985.
- Born R, Bradley D. “Structure and function of visual area MT.”. Annu Rev Neurosci 28: 157-189. PMID 16022593.
- Felleman D, Van Essen D. “Distributed hierarchical processing in the primate cerebral cortex.”. Cereb Cortex 1 (1): 1-47. PMID 1822724.
- Ungerleider L, Desimone R (1986). “Cortical connections of visual area MT in the macaque.”. J Comp Neurol 248 (2): 190-222. doi:10.1002/cne.902480204. PMID 3722458.
- Sincich L, Park K, Wohlgemuth M, Horton J (2004). “Bypassing V1: a direct geniculate input to area MT.”. Nat Neurosci 7 (10): 1123-8. doi:10.1038/nn1318. PMID 15378066.
- Palmer SM, Rosa MG (2006). “A distinct anatomical network of cortical areas for analysis of motion in far peripheral vision.”. Eur J Neurosci 24(8): 2389-405.
- Rodman HR, Gross CG, Albright TD (1989) Afferent basis of visual response properties in area MT of the macaque. I. Effects of striate cortex removal. J Neurosci 9(6):2033-50.
- Dubner R, Zeki S (1971). “Response properties and receptive fields of cells in an anatomically defined region of the superior temporal sulcus in the monkey.”. Brain Res 35 (2): 528-32. doi:10.1016/0006-8993(71)90494-X. PMID 5002708..
- Maunsell J, Van Essen D (1983). “Functional properties of neurons in middle temporal visual area of the macaque monkey. I. Selectivity for stimulus direction, speed, and orientation.”. J Neurophysiol 49 (5): 1127-47. PMID 6864242.
- Hess, Baker, Zihl (1989). “The" motion-blind" patient: low-level spatial and temporal filters”. Journal of Neuroscience 9 (5): 1628-1640.
- Baker, Hess, Zihl (1991). “Residual motion perception in a" motion-blind" patient, assessed with limited-lifetime random dot stimuli”. Journal of Neuroscience 11 (2): 454-461.
- Movshon, J.A., Adelson, E.H., Gizzi, M.S., & Newsome, W.T. (1985). The analysis of moving visual patterns. In: C. Chagas, R. Gattass, & C. Gross (Eds.), Pattern recognition mechanisms (pp. 117-151), Rome: Vatican Press.
- Britten & Van Wezel 1998
- Wilson, H.R., Ferrera, V.P., & Yo, C. (1992). A psychophysically motivated model for two-dimensional motion perception. Vis Neurosci, 9 (1), 79-97.
- Tinsley, C.J., Webb, B.S., Barraclough, N.E., Vincent, C.J., Parker, A., & Derrington, A.M. (2003). The nature of V1 neural responses to 2D moving patterns depends on receptive-field structure in the marmoset monkey. J Neurophysiol, 90 (2), 930-937.
- Pack & Born, 2003
- Albright T (1984). “Direction and orientation selectivity of neurons in visual area MT of the macaque.”. J Neurophysiol 52 (6): 1106-30. PMID 6520628.
- DeAngelis G, Newsome W (1999). “Organization of disparity-selective neurons in macaque area MT.”. J Neurosci 19 (4): 1398-415. PMID 9952417.
関連文献
日本語のオープンアクセス文献
- 佐藤宏道, 勝山成美 「一次視覚野の機能構築とその役割」 日本視能訓練士協会誌 Vol.21 (1993) pp.154-160
- 乾 敏郎 「視覚の計算理論とモジュールの統合」 計測と制御 Vol.33, No.3 (1994) pp.222-226
外部リンク
- 第一次視覚野が壊れると見えなくなる - 中部学院大学 三上章允
- サルやヒトの脳には沢山の視覚野がある - 中部学院大学 三上章允
- ユタ大学のマシュー・スコモレスキー (Matthew Schmolesky) による一次視覚野の解説
- ハーバード大学のデービット・ヒューベルによる視覚野の構造の解説
- 視覚の計算モデリングのシミュレーター
- Area V1 (英語) - スカラーペディア百科事典「一次視覚野」の項目。
- 眼優位性 - 脳科学辞典