遷移 (生物学)
植物群落を中心とした遷移(植生遷移)
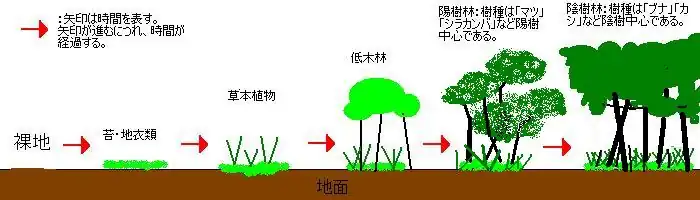
植物が土地で生育することによる、環境形成作用が主な原因となり、時とともに場所の環境が変化して行く現象を植生遷移という。陸上で進行する植生遷移を乾性遷移、水圏で進行する植生遷移を湿性遷移という呼び方もある。
一次遷移
基質に全く生物を含まないところから始まる遷移を一次遷移と言う。
生物やその他の風化の影響を受けていない地球表面というものを考える。 自然界においては、溶岩流や氷河の侵食によって作られた土地、岩盤の上等がそれに当たるが、そのような土地は現代では極めて限られるため、一次遷移が起こりえる土地というのも限定的である。そのため、一次遷移より、後述する二次遷移の方が、自然界では起こりやすい。
- 岩盤のくぼみや裂け目などに、岩石表面そのものが徐々に風化して得られたり、風雨によって運ばれた砂礫が堆積し、土壌を形成する。この土壌の形成の有無が一次遷移と二次遷移の最大の違いである。土壌のないところは保水力がないため、植物は生育できない。よって、この土壌の形成が最初の大きな変化である。
- 次に、苔類や地衣類の胞子が風雨によって運ばれると考えられる。コケ類は岩のくぼみのようなところを中心にはえてくる。やがて、風や雨によって岩の表面は風化して砂礫が生じ、また、苔や地衣類の生育による有機物の蓄積により少しずつ土壌が成熟される。土壌には土壌動物や土壌微生物も当然出現する。
- ある程度の量の土壌が蓄積すれば、草本が侵入することができるようになる。当初は一年性草本が中心になるが、年を追って多年性草本が量を増やし、次第に背の高い草原となる。草の根は苔より深くまで侵入し、砂礫と土壌の層は厚くなる。土壌中には、ミミズなど大型の土壌動物も姿を見せ、陸上には昆虫や鳥も侵入する。
- やがて、木本の侵入が始まる。あらゆる種類の木本植物が進入するが、最初に低木林が形成される。
- 陽樹が優勢の陽樹林が形成される。この段階でも陰樹(後述)の侵入も起こるが、陽樹には最大光合成量が高いものが多く、比較的成長も早いために優勢となりやすい。陽樹が選択的に侵入または生育するわけではなく、あくまでこの環境条件で優勢になりやすいということである。代表的な陽樹にはシラカンバやマツなどがある。陽樹が成長してくると、その下は次第に日陰になるので、草原の植物は勢いを失う。その代わりに日陰であっても成長可能な植物が侵入する。こうして陽樹林ができる。樹木は草本よりも深く根を下ろし、土壌層はさらに厚くなる。土壌にも陸上にも、動物相はさらに豊富になる。
- 森林ができると、その内部は湿度が高く、林床の照度は低くなる。こうなると、陽樹の苗木が生育しにくくなる。その代わり、暗い林床でも成長できる種類の樹木が出てくる。これを陰樹という。陰樹が成長し、森林を構成する樹木になると、しばらくは陽樹と陰樹が交じった森林になるが、陽樹は追加されにくいため、次第に陰樹林となる。陰樹林内では陰樹は生育できるので、見かけ上はこの形の森林がこれより変わることはなくなる。この状態を極相(クライマックス)と言う。土壌層は豊かになり、地上には森林性の草本が次第に生えるようになる。
- 極相林になれば、その後は外見上の大きな変化は少なくなる。しかし、森林性の植物の侵入は時間がかかるし、樹木が大きくなるのも時間がかかる。大径木がなければ生息できない動物もあり、森林が真に成熟するのはその先である。原生林と呼ばれるのは、何百年も森林であったような森である。
- 極相(クライマックス)を迎えた森でも、部分的には常に変化が起こっている。たとえばギャップダイナミクスがその代表的なひとつである。
二次遷移
まず土壌がすでに存在すること、そして基質である土壌に若干の生物、例えば土壌中の種子(埋土種子集団 soil seed bank)・地下茎・土壌動物などを含む場所から始まる遷移を二次遷移という。現代の地球上には、一次遷移が見られる場合というのはほとんどないため、我々が目にすることのできる遷移はおおむねこの二次遷移である。
- 日本で通常見かけるのは、例えば宅地造成のような開けた場所を作った場合、畑地を放棄した場合などである。いずれの場合も、現場に植生はなくとも、土壌があり、土壌中に種子や動植物の死骸などの有機物があるなど、草本が生育する条件は整っている。したがって、この場合、すぐに草本が侵入し、そこから遷移が始まる。数年で多年生草本が繁茂し、ほどなく陽樹の侵入も始まる。そこからは、ほぼ一次遷移と同じ経過を辿ることになる。
- 森林を伐採した場合は、若干異なった形になる。木がなくなっても、草は残っているし、肥沃な土壌もある。また、土壌中にはさまざまな植物の種子が含まれる。それに、木をすべて伐り倒したとしても、株が生きている場合が多く、根元から新芽が伸び始める。かといって、すぐに元の森林に戻る訳ではない。多くの場合、森林性の草本は一時姿を消す。一年目には一年性草本が一面にはえるが、森林性の植物の中でも強い日差しに耐えられる植物が伸び始めるので、翌年にはそれらが地表を覆う。また、森林内では発芽できないでいた陽樹の種子が発芽する。切り株からは元の森林の樹種の新芽が出始めるが、陽樹の芽はそれより早く伸び、一時的に陽樹の多い林になる。やがて陰樹がそれらに取って代わり、元の森林に戻って行く。このように、どの段階で、どのような撹乱が起きたかによって、そこから始まる遷移の状況は若干変わってくる。
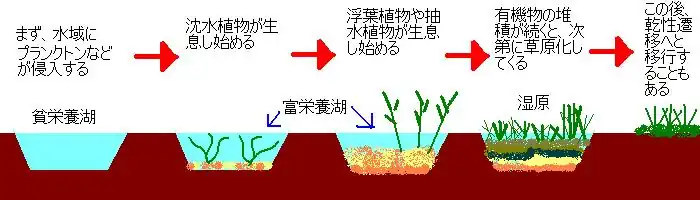
その他の遷移
糞生菌の遷移
動物の糞は、様々な菌群が観察できることが(菌類学徒の間では)よく知られている。糞以外からも出現する菌もあれば、ほとんど糞からのみ出現するものもある。その出現の順番には、ほぼ一定の型があり、糞生菌の遷移と言われる。
通常、最初に出現するのは、接合菌門のケカビ類である。ケカビ、ミズタマカビなどは特に頻繁に出現する。後者は、ほぼ糞からのみ出現する種である。接合菌類の出現は2〜3日目から、1週間くらい続く。ほぼ同じか、少し遅れて小型の不完全菌類が出現する。不完全菌はかなり遅い時期まで出続ける。
1週間目くらいから、1mm以下程度の小型の子実体を作る子嚢菌門の菌が姿を現す。さらに、2〜3週間目くらいにヒトヨタケなど担子菌門(キノコ類)が出現すると、これ以後は次第に糞生菌から通常の土壌菌の群集へと移ってゆく。
この遷移の原因は、以下のようなものだと考えられている。
その他の場合
枯葉や枯れ枝などの場合にも糞生菌とほぼ同様な遷移があることが知られる。新鮮な落ち葉には接合菌が生育し、短時間で胞子形成を行うと姿を消す。子嚢菌はやや遅れて出現し、長時間にわたって生育する。原因は糞生菌の場合とほぼ同じと考えられている。