エピクロエ属
エピクロエ属(エピクロエぞく、Epichloë)とは、子嚢菌門真菌の属の一つである。イネ科植物の茎、葉、花序、および種子の細胞間に生息し、宿主植物と内部共生する。がまの穂病の病原菌でもある。この病害では春の終わりの穂ばらみ期から出穂期に葉鞘にコロニーを形成し、花序の発達と出穂を妨げる。それ以外の、宿主植物の生活環の大部分では病原性を示さない。むしろ、二次代謝産物産生での草食動物による食害の防止、宿主のストレス耐性の向上、生育の促進など、宿主に利益を与える。
| エピクロエ | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 がまの穂病:Epichloë typhina によるナガハグサPoa pratensisの子座(ストロマ) | |||||||||||||||||||||
| 分類 | |||||||||||||||||||||
| |||||||||||||||||||||
| 学名 | |||||||||||||||||||||
| Epichloë (Fr.) Tul. & C. Tul | |||||||||||||||||||||
| タイプ種 | |||||||||||||||||||||
| Epichloë typhina (Fr.) Tul. & C. Tul | |||||||||||||||||||||
| 種 | |||||||||||||||||||||
|
本文参照 |
エピクロエ属は現在、有性世代(テレオモルフ)と無性世代(アナモルフ)の両方を含む。以前、この無性世代はネオティホディウム属に属していた。分子系統学的な研究により、ネオティホディウム属の多くはエピクロエ属に分類することが適当であることが明らかとなり、再分類が行われた。
エピクロエ属は宿主にとって、その生育と生態に重大な影響を与える存在である。多くのイネ科植物、特にオニウシノケグサとライグラスでは、エピクロエ属を含む内生菌に感染した品種が芝草として商品化されている。
分類
1849年、エリーアス・フリースはノムシタケ属(Cordyceps)の亜属としてエピクロエという名称を用いた[1]。彼はこのとき、クリスティアーン・ヘンドリク・ペルズーンが発見したCordyceps typhinaを基準種に設定した[1]。1865年、シャルル・テュラーヌとエドモン・テュラーヌの兄弟は亜属を属に格上げした[2]。これにより、ノムシタケ属だった基準種はEpichloë typhinaとなった。当初はエピクロエ属の種はこの基準種のみだったが、1970から1980年代に、畜産動物の中毒症状の原因菌として牧草から新種が発見された[3]。1998年、Parepichloë属の一部がエピクロエ属に再分類された[4]。2002年、日本特産のEpichloë sasaeと東南アジア産のEpichloë bambusaeが新設されたHeteroepichloë属へ移された[5]。
現在、エピクロエ属の多くは無性および有性生殖の形態両方(それぞれ無性世代、有性世代)を持ち、さらにいくつかの種は無性世代しか持たない。しかし、かつて、両世代を有する種について、無性世代と有性世代は異なる菌種として認識されていた。無性世代はエピクロエ属とは異なる分類群に分類されていた。
1982年、ギャレス・モーガン=ジョーンズ(Gareth Morgan-Jones)とヴァルター・ガムスにより、無性世代はアクレモニウム属のAlbo-lanosa節に再分類された[6]。1996年、Glennと共同研究者は分子系統解析により当時のアクレモニウム属が多系統群であることを知った(通常、多系統群であることが明らかとなった場合、異なる系統群を異なる分類群に分離するため再分類が行われる)。彼らは新しくネオティホディウム属を作り、エピクロエ属に関連する無性世代をこの新属の分類することを提案した[7]。2014年、 Leuchtmannと共同研究者はネオティホディウム属のほとんどがエピクロエ属の無性世代であることを確認し、これらをエピクロエ属に再分類した[3]。なぜなら、国際藻類・菌類・植物命名規約は2011年から無性世代と有性世代で異なる名前を付けないようにし、全ての世代で同一の名前を付けるように求めているためである[3]。
種
2016年現在、34種が登録されている。これとは別に亜種や変種も存在する。それらを下記に示す。アスタリスク*で印を付けられた種は現在のところ無性世代のみが明らかとなっているものである[3]。
- Epichloë amarillans
- Epichloë aotearoae*
- Epichloë australiensis*
- Epichloë baconii
- Epichloë brachyelytri
- Epichloë bromicola (syn. E. yangzii)
- Epichloë cabralii
- Epichloë canadensis*
- Epichloë chisosa*
- Epichloë coenophiala*
- Epichloë danica*
- Epichloë disjuncta*
- Epichloë elymi
- Epichloë festucae
- var. lolii*
- Epichloë funkii*
- Epichloë gansuensis*
- var. inebrians*
- Epichloë glyceriae
- Epichloë guerinii*
- Epichloë hordelymi
- Epichloë liyangensis
- Epichloë melicicola*
- Epichloë mollis*
- Epichloë occultans*
- Epichloë pampeana*
- Epichloë schardlii*
- Epichloë sibirica*
- Epichloë siegelii*
- Epichloë sinica*
- Epichloë sinofestucae*
- Epichloë stromatolonga*
- Epichloë sylvatica
- subsp. pollinensis
- Epichloë tembladerae*
- Epichloë typhina
- var. ammophilae*
- subsp. clarkii
- subsp. poae
- var. aonikenkana
- var. canariensis
- var. huerfana*
- Epichloë uncinata*
生活環
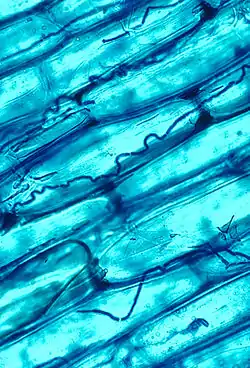
エピクロエ属は、植物と内部共生するよう進化しており、宿主への病原性は全くないか、通常条件では発現しないように制限されている 。この共生関係について最も研究が進んでいるのはイネ科とカヤツリグサ科についてである。エピクロエ属は葉などの地上部組織において、細胞間に(内生成長)もしくは上皮の上か下の表面に(着生成長)感染する[8]。
エピクロエ属は植物だけでなく、ハナバエ科Botanophila属のハエとも共生関係にある。ハエはエピクロエ属の子座の菌糸に卵を産み、幼虫はこの菌糸を餌とする。その対価として、成虫はエピクロエ属の子嚢胞子を体に着けて植物個体間の水平伝播を媒介する[9]。
進化
エピクロエ属が古くから植物に感染していたことは多くの証拠から明らかとなっている。今日のイネ科イチゴツナギ亜科が誕生した時には共生関係は成立していた。他の真菌属と比べ、植物宿主との共拡散は優勢であったことが系統樹で示されている[10]。
内生真菌の成長は宿主植物内で厳密に制御されている。例えば、外部環境とは異なり宿主植物内では菌糸は殆ど分岐しない[11][12]。また、植物の葉の成長と、菌糸の伸長拡大は同期している。この同期性は他の真菌種には見られない[13]。また、同期性にはNADPHオキシダーゼ複合体による活性酸素の生成機構が必須である。この酵素の遺伝子を欠失させたEpichloë festucaeは宿主植物で無秩序に成長し、宿主の枯死すら招く[14][15]。
エピクロエ属の無性種の由来は、一般的に、少なくとも異なるエピクロエ属2種の交配であることが明らかとなっている[16][17]。
化学物質の産生
エピクロエ属は二次代謝産物として、草食昆虫および哺乳動物に対する毒素、もしくは摂食抑制剤を産生する[18]。
エピクロエ属は、草食動物に対して様々な生理活性を有する化学物質を産生する[19][20]。麦角アルカロイド類はエピクロエ属の代表的な毒素であり、エルゴリン骨格のアルカロイド類である[21]。
麦角アルカロイド類のうち、宿主植物の体内で最も多く存在するのはエルゴバリンである。エルゴバリンは、L-プロリン、L-アラニン、およびL-バリンと結合した二環トリペプチドと結合したエルゴリン部分を有する。この部分の合成に関わる以下の二つの遺伝子はエルゴバリンの鍵遺伝子である。dmaWはジメチルアリルトリプトファン合成酵素を、lpsAは非リボソームペプチド合成酵素をそれぞれコードしている[21]。
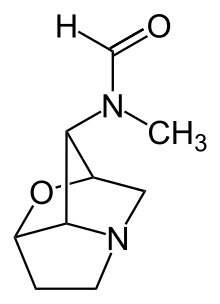
そのほか、エピクロエ属が産生する二次代謝産物を以下に示す。ロリトレムBなどのインドールジテルペン類はいくつかの酵素、例えばプレニル基転移酵素やモノオキシゲナーゼに対して活性を持つ[22] 。エルゴリンとインドールジテルペン類のどちらも草食哺乳類一般と一部の昆虫に対して有毒である[19] 。ペラミンは、L-アルギニンとピロリチジン-5-カルボン酸から生合成されるピロリチジンアルカロイドの一種である[23] 。昆虫の摂食を抑制する。ロリンアルカロイド類は、1-アミノピロリジジンで2位炭素と7位炭素に酸素が結合したものである[24] 。L-プロリンとL-ホモセリンから生合成され[25] 。ロリンは、ニコチンに匹敵する耐昆虫の殺傷と摂食抑制効果を持つ[24] 。エピクロエ属を食した昆虫において、特に若い組織でロリンは蓄積し[26] 、障害を受ける[27] 。
例外はあるが多くの場合、エピクロエ属は麦角アルカロイド類、インドールイソプレノイド類、ロリンアルカロイド類の3種を産生する[19] 。Epichloë uncinataの感染によるロリンは甲虫Heteronychus aratorからイネ科Festulolium属を防御する[28]。
宿主への影響
エピクロエ属の繁殖と生存は宿主のそれらと密接に関連しており、この真菌の垂直伝播は相利共生へと進化した、あるいはしていくと考えられている[29]。事実、エピクロエ属は宿主に成長促進、旱魃耐性、草食動物や病原体への抵抗性などの利益を与えている[30][31]。エピクロエ属は一般的に野生や栽培下の植物に対して相利共生であると認識されている[32][33]。しかし、特に土壌栄養素が貧しい条件で、ときに害をなすこともある[34]。
生態系への影響
エピクロエ属を保有する草本種は比較的多く、保有草本がいる環境も多様かつ多数である。エピクロエ属保有による利益や病害が自然や農業環境にどのような影響を及ぼしているのか完全には明らかになっていない[30][35]。エピクロエ属保有草本の増加と草食動物の摂食に何らかの影響があることが示唆されている[19][23]。草食動物による攻撃や環境条件によって内生真菌は複雑な相互作用を示す[36]。エピクロエ属は、草食動物に対して有毒な化学物質を産生する[37][38] 。植物の生育や生理に直接影響を及ぼす 。
脚注
- Fries, E.M. (1849) (Latin). Summa vegetabilium Scandinaviae. Stockholm, Leipzig: Bonnier. pp. 572
- Tulasne, L.R.; Tulasne, C. (1865) (Latin). Selecta Fungorum Carpologia: Nectriei – Phacidiei – Pezizei. 3
- Leuchtmann, A.; Bacon, C. W.; Schardl, C. L.; White, J. F.; Tadych, M. (2014). “Nomenclatural realignment of Neotyphodium species with genus Epichloë” (PDF). Mycologia 106 (2): 202–215. doi:10.3852/13-251. ISSN 0027-5514. PMID 24459125.
- White, J.F.; Reddy, P.V. (1998). “Examination of Structure and Molecular Phylogenetic Relationships of Some Graminicolous Symbionts in Genera Epichloe and Parepichloe”. Mycologia 90 (2): 226. doi:10.2307/3761298. ISSN 0027-5514. JSTOR 3761298.
- Heteroepichloë , gen. nov. (Clavicipitaceae; Ascomycotina) on bamboo plants in East Asia
- Morgan-Jones, G.; Gams, W. (1982). “Notes on hyphomycetes. XLI. An endophyte of Festuca arundinacea and the anamorph of Epichloe typhina, new taxa in one of two new sections of Acremonium”. Mycotaxon 15: 311–318. ISSN 0093-4666.
- “Molecular phylogeny of Acremonium and its taxonomic implications”. Mycologia (Mycological Society of America) 88 (3): 369–383. (1996). doi:10.2307/3760878. JSTOR 3760878.
- “Production of loline alkaloids by the grass endophyte, Neotyphodium uncinatum, in defined media”. Phytochemistry 58 (3): 395–401. (2001). doi:10.1016/S0031-9422(01)00272-2. PMID 11557071.
- Górzyńska, K. (2010). “An unusual Botanophila–Epichloë association in a population of orchardgrass (Dactylis glomerata) in Poland”. Journal of Natural History 44 (45-46): 2817–24. doi:10.1111/j.1570-7458.2006.00518.x.
- “A novel test for host-symbiont codivergence indicates ancient origin of fungal endophytes in grasses.”. Syst Biol. 57 (3): 483–498. (2008). doi:10.1080/10635150802172184. PMID 18570040.
- “In planta regulation of extension of an endophytic fungus and maintenance of high metabolic rates in its mycelium in the absence of apical extension”. Appl. Environ. Microbiol. 67 (12): 5377–5383. (2001). doi:10.1128/AEM.67.12.5377-5383.2001. PMC 93319. PMID 11722882.
- “Growth of Epichloë/Neotyphodium and p-endophytes in leaves of Lolium and Festuca grasses”. Mycol. Res. 96: 93–106. (2002). doi:10.1017/S095375620100510X.
- “Epichloë endophytes grow by intercalary hyphal extension in elongating grass leaves”. Fungal Genet. Biol. 45 (2): 84–93. (2008). doi:10.1016/j.fgb.2007.07.013. PMID 17919950.
- “Reactive oxygen species play a role in regulating a fungus-perennial ryegrass mutualistic interaction”. Plant Cell 18 (4): 1052–1066. (2006). doi:10.1105/tpc.105.039263. PMC 1425850. PMID 16517760.
- “A p67Phox-like regulator is recruited to control hyphal branching in a fungal-grass mutualistic symbiosis”. Plant Cell 18 (10): 2807–2821. (2006). doi:10.1105/tpc.106.046169. PMC 1626622. PMID 17041146.
- “Evolutionary diversification of fungal endophytes of tall fescue grass by hybridization with Epichloë species”. Proc. Natl. Acad. Sci. USA 91 (7): 2542–2546. (1994). doi:10.1073/pnas.91.7.2542. PMC 43405. PMID 8172623.
- “Prevalence of interspecific hybrids amongst asexual fungal endophytes of grasses”. Molec Ecol 13 (6): 1455–1467. (2004). doi:10.1111/j.1365-294X.2004.02138.x. PMID 15140090.
- Roberts CA, West CP, Spiers DE, eds (2005). Neotyphodium in Cool-Season Grasses. Blackwell. ISBN 978-0-8138-0189-6
- “Bioprotective Alkaloids of Grass-Fungal Endophyte Symbioses”. Plant Physiol. 114 (1): 1–7. (1997). doi:10.1104/pp.114.1.1. PMC 158272. PMID 12223685.
- Scott B (2001). “Epichloë endophytes: fungal symbionts of grasses”. Curr. Opin. Microbiol. 4 (4): 393–398. doi:10.1016/S1369-5274(00)00224-1. PMID 11495800.
- “Ergot alkaloids – biology and molecular biology”. The Alkaloids: Chemistry and Biology 63: 45–86. (2006). doi:10.1016/S1099-4831(06)63002-2. PMID 17133714.
- “A complex gene cluster for indole-diterpene biosynthesis in the grass endophyte Neotyphodium lolii”. Fungal Genet Biol 43 (10): 679–693. (2006). doi:10.1016/j.fgb.2006.04.004. PMID 16765617.
- Tanaka A, Tapper BA; Popay A, Parker; EJ, Scott B (2005). “A symbiosis expressed non-ribosomal peptide synthetase from a mutualistic fungal endophyte of perennial ryegrass confers protection to the symbiotum from insect herbivory”. Mol. Microbiol. 57 (4): 1036–1050. doi:10.1111/j.1365-2958.2005.04747.x. PMID 16091042.
- “Loline alkaloids: currencies of mutualism”. Phytochemistry 68 (7): 980–996. (2007). doi:10.1016/j.phytochem.2007.01.010. PMID 17346759.
- “Biosynthetic precursors of fungal pyrrolizidines, the loline alkaloids”. Chembiochem 6 (6): 1016–1022. (2005). doi:10.1002/cbic.200400327. PMID 15861432.
- Zhang, DX, Nagabhyru, P, Schardl CL (2009). “Regulation of a chemical defense against herbivory produced by symbiotic fungi in grass plants”. Plant Physiology 150 (2): 1072–1082. doi:10.1104/pp.109.138222. PMC 2689992. PMID 19403726.
- “Stroma-forming endophyte Epichloe glyceriae provides wound-inducible herbivore resistance to its grass host”. Oikos 117: 629–633. (2008). doi:10.1111/j.0030-1299.2008.16483.x.
- Barker GM; Patchett BJ; Cameron NE (2014). “Epichloë uncinata infection and loline content afford Festulolium grasses protection from black beetle (Heteronychus arator). Landcare Research, Hamilton, New Zealand & Cropmark Seeds Ltd, Christchurch, New Zealand 20 Dec 2014.”. New Zealand Journal of Agricultural Research 58: 35–56. doi:10.1080/00288233.2014.978480.
- Ewald PW (1987). “Transmission modes and evolution of the parasitism-mutualism continuum”. Ann NY Acad Sci 503: 295–306. doi:10.1111/j.1749-6632.1987.tb40616.x. PMID 3304078.
- “Symbioses of grasses with seedborne fungal endophytes”. Annu Rev Plant Biol 55: 315–340. (2004). doi:10.1146/annurev.arplant.55.031903.141735. PMID 15377223.
- “Adaptations of endophyte-infected cool-season grasses to environmental stresses: mechanisms of drought and mineral stress tolerance”. Crop Sci. 40: 923–940. (2000). doi:10.2135/cropsci2000.404923x.
- “Herbivores cause a rapid increase in hereditary symbiosis and alter plant community composition”. Proc. Natl. Acad. Sci. USA 102 (35): 12465–12470. (2005). doi:10.1073/pnas.0503059102. PMC 1194913. PMID 16116093.
- “Herbivory mediates grass-endophyte relationships”. Ecology 88 (11): 2752–2757. (2007). doi:10.1890/06-1958.1. PMID 18051643.
- “The persistence of vertically transmitted fungi in grass metapopulations”. Proc Biol Sci 269 (1498): 1397–1403. (2002). doi:10.1098/rspb.2002.2006. PMC 1691040. PMID 12079664.
- “Model systems in ecology: dissecting the endophyte-grass literature”. Trends Plant Sci 11 (9): 428–433. (2006). doi:10.1016/j.tplants.2006.07.001. PMID 16890473.
- “Variation in the abundance of fungal endophytes in fescue grasses along altitudinal and grazing gradients”. Ecography 3: 422–430. (2007). doi:10.1111/j.0906-7590.2007.05027.x.
- Hahn H, McManus MT, Warnstorff K, Monahan BJ, Young CA, Davies E, Tapper BA, Scott, B (2007). “Neotyphodium fungal endophytes confer physiological protection to perennial ryegrass (Lolium perenne L.) subjected to a water deficit”. Env. Exp. Bot. 63: 183–199. doi:10.1016/j.envexpbot.2007.10.021.
- Hunt MG.; Rasmussen S; Newton PCD; Parsons AJ; Newman JA (2005). “Near-term impacts of elevated CO2, nitrogen and fungal endophyte-infection on Lolium perenne L. growth, chemical composition and alkaloid production”. Plant Cell Environ. 28: 1345–1354. doi:10.1111/j.1365-3040.2005.01367.x.
外部リンク
- エピクロエ属 in Index Fungorum
- エピクロエ属 in MycoBank.