アマノリ
アマノリ(甘海苔)は、紅藻のウシケノリ綱ウシケノリ目ウシケノリ科に属する海藻のうち、膜状の体をもつ種の総称であり、アサクサノリやスサビノリ、ウップルイノリなどが含まれる。このような膜状の体は葉状体とよばれ、1層 (まれに2層) の細胞層からなり、生活環における配偶体 (ゲノムを1セットもち配偶子をつくる体) である。葉状体が放出した果胞子は発芽して微小な胞子体 (ゲノムを2セットもち胞子をつくる体) になり、ふつう貝殻などに穿孔する (糸状体とよばれる)。日本など東アジアでは古くから食用とされ、現在では大規模に養殖されている。またイギリスやハワイなど世界各地でも採取され食材とされている。
| アマノリ類 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||
| 分類 | |||||||||||||||||||||
| |||||||||||||||||||||
| 英名 | |||||||||||||||||||||
| foliose Bangiales, nori, laver | |||||||||||||||||||||
| 下位分類 | |||||||||||||||||||||
|
日本におけるアマノリ類に対する漢字表記としては、古くは漢名の「神仙菜」や「紫菜」が用いられていた[1]。鎌倉時代以降は「甘海苔」が使われるようになり、さらに江戸時代には単に「海苔」と記すようになった[1]。ただし公文書では大宝律令 (701年) 以降、明治時代まで「紫菜」が使われていた[1]。また「海苔」(ノリ) という語は、アオノリやカワノリ、ハバノリ、スイゼンジノリなど遠縁の食用藻類を含む広い意味で用いられることも多い。
2020年現在世界中で160種以上が知られ、日本からは約30種が報告されている。2011年まではこのような藻類は全てポルフィラ属 (Porphyra;和名ではアマノリ属とよばれていた) にまとめられることが多かった。しかし分子系統学的研究の結果、この意味でのポルフィラ属は単系統群ではないことが示され (多列糸状であるウシケノリ類と分けられない)、多数の属に細分されることになった。2020年現在では、アマノリ類はおよそ14属に分けられている。この過程で和名のアマノリ属に充てる学名は Porphyra、Pyropia、Neopyropia と変遷している[2]。
特徴
形態と生活環

Clip
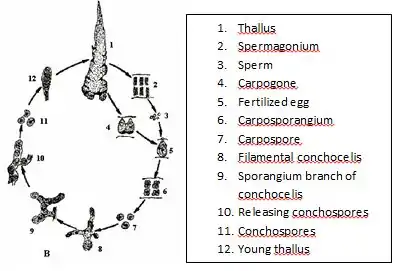
基本的に巨視的な配偶体 (葉状体) と微細な胞子体 (糸状体) の間で世代交代を行う[3][4][5][6] (右図1b)。生活環における世代交代の時期は種によって異なり (#生態参照)、水温や日長で調整されている。
配偶体は薄い葉状 (膜状) の体をもち、葉状体ともよばれる[3][7] (右図1a)。葉状体の形は円形から披針形、長さ数 cm から 30 cm ほどのものが多いが、まれに 1 m 以上になるものもいる[7][8]。葉状体はふつう1層の細胞層からなり厚さはふつう 20-70 μmほどであるが、ベニタサ属は2層の細胞層からなり厚さ 150 μmに達するものもいる[7]。葉状体は仮根で岩や杭、貝殻や他の海藻などの基質に着生している[9]。葉状体の細胞中央には、1個の星形の葉緑体が存在する (一部の種は2個の星形葉緑体が細胞中にならんでいる)[7][10]。星形葉緑体の中央にはピレノイドが存在する。周縁チラコイド (色素体膜の内側に位置し、他のチラコイドを囲んでいるチラコイド) をもたない。細胞壁の繊維多糖としてキシランやマンナンをもつ[11]。配偶体の一部の細胞は造果器 (carpogonium) または造精器 (精子嚢, antheridium, spermatangium) になる。同じ個体に造果器と造精器をつけるもの (雌雄同株) と、どちらか一方のみを付けるもの (雌雄異株) がある[7]。雌雄同株の場合、雌雄が左右など明らかに分離して形成される種と、混在して形成される種がある[7]。造果器は雌性配偶子嚢であり、葉状体の片側または両側に突起 (受精突起, trichogyne, protrichogyne) をもつが、不明瞭なこともある[7][12]。造精器は細胞分裂をして多数の不動精子 (精子, spermatium) を形成する。不動精子は放出され、造果器に付着して受精する。受精した造果器は細胞分裂を行い、果胞子 [接合胞子 (zygotospore) ともよばれる[12]] を形成する。果胞子は直径 15 μmほど[3]。不動精子や果胞子の形成において、縦横高さの細胞数がそれぞれ種によってほぼ決まっており、分類形質とされる[7]。
果胞子は貝殻など石灰質基質上で発芽し、微小な分枝糸状体である胞子体 (糸状体) になる[3]。糸状体は細くふつう直径 2-5 μmほどであり、貝殻中では直線的に伸び、対生状に分枝する[3][13][14]。糸状体は貝殻など石灰質基質中に穿孔するが、ゲノム情報からは炭酸脱水酵素を分泌することによって炭酸カルシウムを溶かしていることが示唆されている[15]。おそらくこの反応によって、糸状体は光合成のための二酸化炭素を得ている[15]。また石灰質基質に穿孔していることで、糸状体は紫外線を含む強光や乾燥から保護されている[16]。この糸状体は古くは別の藻類と考えられ、Conchocelisという属名が与えられていたため、現在ではコンコセリス期とよばれることもある[4]。葉状体は特定の分裂細胞をもたないが、糸状体は頂端に分裂細胞をもつ先端成長を行う。糸状体を構成する細胞は1個から数個の側膜性葉緑体をもち、葉緑体中には1個から2個のピレノイドが存在する[10]。また葉緑体が周縁チラコイドをもつ点でも葉状体とは異なる。糸状体の細胞壁を構成する繊維多糖はセルロースである[11]。葉状体の細胞間にはピットプラグは存在しないが、糸状体の細胞間には存在する[17]。糸状体は殻胞子嚢を形成し、殻胞子 (conchospore) を放出する。殻胞子は直径 10-15 μmほどであり、岩などの基質に付着して発芽し、葉状体となる[3][13][14]。この発芽・成長の過程で減数分裂が起こると考えられている[3]。カイガラアマノリは殻胞子を形成せず、貝殻中の糸状体から直接、葉状体が形成される (そのため葉状体が貝殻から生じている)[3]。
種によっては、葉状体 (特に若い時期) の一部の細胞がそのまま原胞子 (archeospore; 単胞子または中性胞子とよばれることも多い[3]) となって散布され、再び葉状体を形成することで無性生殖を行う[3][6][12]。葉状体の一部の細胞が受精を経ずに細胞分裂をして形成された胞子のうち、葉状体へと発生するものは中性胞子 (neutral spore)、糸状体へと発生するものは無配胞子 (agamospore) とよばれる[6][12]。また糸状体が原胞子によって再び糸状体を形成する無性生殖を行う例も知れられている[6][12]。
色素
葉緑体はクロロフィル a を含み、カロテノイドとしてゼアキサンチン、ルテイン、β-カロテン、α-カロテンをもつ[18]。色素タンパク質としてアロフィコシアニン、フィコシアニン、フィコエリスリンが存在し、チラコイド上でフィコビリソームを形成している。アマノリの色は、主にクロロフィルの緑色とフィコエリスリンの赤色の組み合わせによる。焼き海苔にするとフィコエリスリンなどが壊れるためクロロフィルの緑色が目立つようになり、また板海苔を高温多湿下におくとクロロフィルが壊れて相対的にフィコエリスリンの赤味が目立つようになる[19]。
生態
_-_geograph.org.uk_-_1247737.jpg.webp)
世界中の寒帯から熱帯までアマノリ類のさまざまな種が生育しており、日本周辺では寒帯性の種 (チシマクロノリ、ベニタサなど)、亜寒帯性の種 (マクレアマノリ、オオノノリなど)、亜寒帯から温帯性の種 (スサビノリ、ウップルイノリなど)、温帯性の種 (クロノリ、マルバアサクサノリなど)、温帯から亜熱帯性の種 (マルバアマノリなど) が見られる[5][7]。
アマノリ類の葉状体は潮間帯の上部に見られるものが多いが (右図1c)、潮下帯 (漸深帯) に生育する例もある[7][20][21]。波の強い外海に面した環境に生育する種 (ウップルイノリなど)[22] から、内湾や河口の干潟のような波静かな環境に生育する種 (アサクサノリなど)[23] がいる[7]。ふつう岩に着生しているが、貝殻やヨシの茎、他の海藻に付着している例もある[23][24][25][26]。葉状体に比べて糸状体は乾燥や強光に弱く、主に潮下帯 (漸深帯) に生育しているものと考えられている[27]。
日本産の種の多くは、秋から春にかけて葉状体 (配偶体) が出現し、夏期は糸状体 (胞子体) として過ごす[3]。ただし葉状体が夏にも残る例がある (マルバアサクサノリなど)[28]。またこれとは逆に葉状体が夏に、糸状体が冬に見られる種もいる (チシマクロノリなど)[29]。
人間との関わり
日本では、アマノリ類は古くから食用として利用されてきた。江戸時代までは野生のアマノリを採取していたが (右図2a)、延宝年間 (1673-1681年) 頃にアマノリが着生しやすい枝や竹 (ひび、そだひび) を遠浅の海に立ててアマノリを養殖する技術が確立され、身近な食材となっていった。その後長い間、アマノリの採苗 (胞子をひびに付ける作業) は自然まかせであったが、1949年にアマノリの生活環が明らかになり、それをもとに人工的にアマノリを採苗する技術が開発された。人工採苗によって特定のアマノリを養殖することが可能になり、2020年現在ではアマノリ養殖のほとんどがスサビノリ (特にその品種であるナラワスサビノリやそれに由来する養殖品種) を用いている。養殖されたアマノリはふつう板海苔に加工され、寿司やおにぎりなどさまざまな食品に利用されている。日本以外でも、世界各地でアマノリは食用とされる。
歴史

_ou_se_fixeront_les_spores_de_l'Asaksamori_on_(Porphyra.jpeg.webp)





日本における文献上のアマノリの初出は『大宝律令』(701年) であり、租税として指定している8種類の海藻の中の1つとしてアマノリ (紫菜) が記されている (他にテングサ、ミル、ワカメ、アラメなど)[30]。その施行日である大宝2年1月1日を西暦に換算すると702年2月6日になるため、全国海苔貝類漁業協同組合連合会が毎年2月6日を「海苔の日」に定めている[31][32]。大宝律令に続く『養老律令』(720年頃) でも、租庸調の調の1品としてアマノリ (紫菜) が記されている。この中に記されている海藻の中では、アマノリが最も価値が高いものとされている[33]。平城京や長岡京跡からは、「紫菜」が記された木簡が出土している[34][35]。同じく8世紀頃の『常陸国風土記』や『出雲国風土記』、『正倉院文書』にも、アマノリの記述がある[30][36]。
『延喜式』(927年完成) では、6カ国 (常陸、志摩、出雲、石見、隠岐、土佐) がアマノリ類の貢納国に指定されている[33][37]。この時代でもアマノリ類は高級品として扱われており、官吏のうち五位以上の者 (貴族) だけにアマノリが支給されていた。また『延喜式』には、寺社の儀式でもアマノリ類が広く使われていたことが記されている[33]。『延喜式』にはアマノリ類を用いた料理として、海菜 (佃煮と考えられている) と汁物 (味噌汁) が記されている[38]。
やがてアマノリの産地として、特定の産地が知られるようになる[39]。出雲国のアマノリは既に『出雲国風土記』(733年完成) に記されており、また十六島 (うっぷるい) にある紫菜島神社 (紫菜島社、のりしまのやしろ) についても記述されている[34]。現在では、十六島はウップルイノリの産地として知られている。『新猿楽記』(11世紀) にも「出雲の浦の甘海苔」の記述があり、1399年には「十六島」の名が献立の記録として残されている[39]。また1186年以降、源頼朝が朝廷へ「伊豆のり」を4回献上したことが記録されている[39]。13世紀には身延山の日蓮に安房からアマノリが届けられ、安房出身である日蓮が故郷を偲ぶ文を残している[39]。
室町時代の『庭訓往来』などには、茶の子 (茶の湯における軽食) や懐石料理としてのアマノリの料理方が記されている[40]。上流階級にとってアマノリは比較的ふつうの食材であり、室町期から江戸初期にかけての4人の公卿の日記ではアマノリの記録が85回見られる (他にコンブ277回、ミル225回、アオノリ84回、モズク62回、ワカメ56回、アワビ359回、ハマグリ146回、カキ42回など)[40]。
『毛吹草』(1645年) は諸国の名産品を挙げており、その中でアマノリ類の産地として、武蔵 (品川苔)、下総 (葛西苔)、安房 (小湊苔)、伊勢、出雲 (十六島苔)、備前 (藤戸苔)、長門 (向津奥苔) が記されている[39]。また同書には、「浅草海苔」の名も初めて見られる。東京湾 (江戸湾) でもおそらく全国各地と同様に古くからアマノリが利用されていたが、江戸という大消費地を背景としていたため、東京湾におけるアマノリの利用は大きく発展することになる。浅草には海苔商が集まり[41]、「浅草海苔」はアマノリの代名詞になっていく。
養殖の始まり
江戸時代の品川周辺などでは漁が行われており、簀立 (すだて;海中に簀で区画を設置して干潮時に逃げおくれた魚を捕らえる仕掛け) や生け簀 (いけす) のために浅海に杭や竹、枝が立てられていた。このようなものにアマノリ (特にアサクサノリ) が着生し、これを採取することが養殖の始まりであったと考えられている[42]。このように自然に着生したアマノリを養殖することは延宝年間 (1673-1681年) 頃に始まったと考えられ[5][43]、また享保2年 (1717年) に品川沖でノリ養殖のために「ひび」(篊、アマノリを着生させるために設置される構造) が立てられた記録がある[42]。ひびの材料としては、大森ではナラ、ケヤキ、クリなどの枝、品川では竹が使われ[42]、このようなひびは「そだひび」ともよばれる[5][42]。これらを 1本ずつまたは数本束にし、端を尖らせて遠浅の海に挿して設置した[42][43] (右図2b)。延享の頃 (1744-1748年) の規格では、40-50 x 2 m ほどの区画 (柵とよばれる) にこのようなひびを40-80本立て、柵と柵の間には海水の流れをよくする流路 (どぶ通しとよばれる) をあけていた[42]。
採取されたアマノリは生品として利用される以外は、そのまま乾燥させたり簀子の上で押し広げて乾燥させていた。やがて享保 (1716-1732) の頃から、アマノリを刻んで漉くことで板海苔がつくられるようになった[42] (右図2d, e, f)。当時の浅草は漉返紙 (浅草紙、再生紙) の産地としても知られており、これが板海苔の開発と関連していたと考えられている[42]。
このようにアマノリ養殖は東京湾で始まったが、養殖技術は次第に日本各地に広まっていった[5][42][44]。安芸国広島湾では、寛保年間 (1741-1743) という比較的早い時期にノリ養殖が始まった[5] (右図2c)。江戸末期までに、陸中国気仙沼、磐城国松川湾、上総国上総湾、安房国鏡カ浦、駿河国三保村、遠江国舞阪宿、尾張国木曽川尻、紀伊国和歌村などでもアマノリ養殖が始まった[5][44]。さらに明治16年 (1883年) に移植の方法が確立され、採苗する場所 (タネ場) と育成する場所を分けることが可能になった[43][45]。その結果、福島で採苗し東京へ移植するなど遠距離の移植も行われるようになった。このような移植方の確立によってさまざまな場所でのアマノリ養殖が可能になり、アマノリ養殖を行う地域が急速に拡大した。
やがて大正から昭和にかけて、「ひび」に大きな変化が起こった[43]。それまでは江戸時代以来のひび立て式 (株立式) が使われていたが、移植法の確立とともに明治後半からノリ養殖業は急速に普及していった。その結果、ひび材が高騰し、またあかぐされ病 (卵菌のフハイカビ属による) の流行から密植が問題視されるようになった。このようなことを背景として網ひびが開発され、またこれを水平に設置することが始まった。このように養殖法には大きな変化が起こったが、アマノリの胞子をひびに付ける作業 (採苗) は自然任せであったため失敗することも多く、アマノリは「運草」ともよばれた[5]。
養殖技術の発展
それまでは、春に葉状体が消失してから秋に再び葉状体が発生するまでの間、アマノリがどのようにして生きているのかは不明であった。しかし1949年、イギリスのキャスリーン・メアリー・ドリュー=ベーカー (Kathleen Mary Drew-Baker) によってアマノリの生活環が明らかにされた[46]。これによって初めて、アマノリ類が夏の間は微小な糸状体 (別の生物として Conchocelis という属名が付けられていたため、現在では糸状体はコンコセリス期ともよばれる[4]) として貝殻の中で生活していることが示された。このことはウェールズなどで食用とされる Porphyra umbilicalis において発見されたが、その後アサクサノリやスサビノリにおいても同様な生活環が確認された[3]。このようなアマノリの生活環の解明により、それまでは自然任せであったものが、特定の種を人工的に採苗することが可能になった。そのため、ドリュー博士は「ノリ養殖の母」とよばれ、熊本県宇土市住吉町の住吉神社境内に顕彰碑が建てられている[5] (右図2g)。その結果、1960年から1970年頃に急速に人工採苗が行われるようになった[5][47][48][49]。さらに1970年頃には冷蔵網技術 および浮流し養殖が普及し、アマノリ養殖は大きな変化を遂げた[5][49]。
またこの頃、養殖種も大きく変化した。1957年頃に東京湾の養殖で見られた種としてはアサクサノリの他にスサビノリ、コスジノリ、マルバアサクサノリ、"チシマクロノリ"[注 1] が報告されていた[50]。このうちアサクサノリは数年間で急速に減少しており (芽いたみ病による減少がきっかけであったことが示唆されている)、代わりにコスジノリやスサビノリが増えていた[50]。他の地域でも同様な種とともに、マルバアマノリ、ツクシアマノリが混在していたことが報告されている[51][52]。人工採苗が一般的になると、病害に強いことや増殖がよいこと、板海苔にしたときに色艶がよいことから、スサビノリが広く使われるようになった。特に1960年代後半に見つかったスサビノリの品種であるナラワスサビノリは成長がよいため主な養殖アマノリとなり、1990年代には養殖アマノリの99%はナラワスサビノリであるとされた[27]。
またアマノリ養殖の中心地も変化していった。江戸時代以来アマノリ養殖の中心地は東京湾であり、第二次世界大戦前には東京湾におけるアマノリ生産は全国の60%を占めていた。しかし他地域の生産量が増加し、1955年頃には全国の30%、2010年頃には4% (ほぼ千葉県のみ) にまで低下している[53][54][55]。また1950年代後半以降の高度経済成長期において東京湾の湾奥部では沿岸開発と水質汚濁が進み、1962年、東京都で沿岸漁業権が全面放棄され、アマノリ養殖も終了した[56]。2020年現在では、日本におけるアマノリ養殖の中心地は瀬戸内海や有明海となっている (下記参照)。
現在の養殖

初期の頃には、アマノリの葉状体をカキ殻に巻き付けておくことで葉状体が放出する果胞子をカキ殻に付着させていたが、現在ではフリー糸状体を用いてカキ殻に糸状体を接種する[5]。フリー糸状体とは、ふつう貝殻に穿孔して育つ糸状体 (胞子体) を、貝殻無しに特殊な培地中で育てたものである[5]。研究所や試験場では、さまざまな栽培品種がフリー糸状体の形で保存されている。まずフリー糸状体を細断し、これをカキ殻に植え付けたものを種苗とする[5][27]。カキ殻中に穿孔した糸状体 (貝殻糸状体) は水槽で培養され、水換えや温度管理によって成長が調整されながら夏を超す。
温度管理によって貝殻糸状体に殻胞子を形成させ、これを網に着生させる (採苗、種付け、殻胞子付け)。東北地方では8月中旬、それ以外では9-10月に採苗を始める[27]。主な方法として野外採苗と室内採苗の2つがある。野外採苗では、糸状体を伴う貝殻をネットやビニール袋に入れ、これを網と一緒に海に張り、海中で採苗する[5]。また海にシートを広げ、その上に貝殻と網を置いて採苗する方法もある[27]。室内採苗では、糸状体を伴う貝殻を入れた水槽中に網をくぐらせることで採苗する[5][27]。
採苗した網 (1.5 x 18 m) は海に張って育苗する[27]。これをそのまま養殖するものは秋芽網とよばれる[5][27]。一方、1ヶ月ほど育苗して長さ2-3 cmに成長した状態で半乾燥・冷凍保存 (-20℃から-30℃) するものは冷蔵網とよばれる[5][27]。冷蔵網は300日以上保存可能であり、1回目の収穫後にこれを設置することで漁期の延長、複数回の収穫が可能になる[5]。また冷蔵網は病原生物や雑藻の除去にも利用できる[5]。
養殖方法には大きく分けて支柱式と浮流し式がある。支柱式では、比較的浅い海で竹やグラスファイバー製の支柱を立て、そこに網を水平に張って固定する[5][27] (右図2h)。また支柱に沿って浮き沈み可能な網を設置する方式もある[5]。浮流し式では、比較的水深の深い場所 (水深 10–20 m 以深) で碇などで固定して水面に浮かべた枠に水平に網を張る[5][27]。浮流し式は養殖漁場設営の制限を受けず、管理しやすいため、大規模な養殖に適している[5]。
養殖の際には、網を一定期間海上に出して干上がらせる (干出) ことがある。干出を与えることによって、アマノリ以外の海藻 (雑藻) の混生や微細藻の付着を防ぎ、また無性生殖のための原胞子 (単胞子ともよばれる) を多く放出させることができる[5]。支柱式の場合は、網を張る高さを調節することで自然な潮の干満によって干出させることが出来る[5][27]。浮流し式の場合は人工干出式浮上筏を用いることもあるが、干出を与えず、雑藻が混入した場合は網を冷蔵処理して雑藻を除去する場合もある (上記)[27]。また雑藻や病原生物を除去するためにクエン酸やリンゴ酸を主成分とする酸性溶液で葉状体を洗浄することがあり、酸処理とよばれる[5]。
採苗から30-40日ほどで葉状体は 10–15 cm になり、摘採される[27]。1回の摘採によって、1枚の網から 20 kg (湿重) ほど得られる。摘採された網はそのまま養殖され、成長したら再び摘採されることを繰り返し、1つの網はふつう7-10日間隔で計4-5回摘採される[27]。摘採には、回転式の摘採機を備えた船が用いられる。摘採時期は、スサビノリで11月上旬から4月、アサクサノリやコスジノリで11月から2月、マルバアサクサノリで12月から4月、ウップルイノリで11月から12月である (あった)[5]。
平成29年度の日本のアマノリ類年間生産量は約30万トン、70-80億枚に達する[57][58][59]。主な生産地は松島湾、東京湾、伊勢湾、瀬戸内海、有明海などであり、特に瀬戸内海や有明海の生産量が多い[27][60]。
上記のように、2020年現在では主にスサビノリ (特にナラワスサビノリ) が養殖に用いられている[8][27]。しかし養殖種の多様化や付加価値のある種の養殖の研究も進められており[61][62]、アサクサノリの養殖復活や地域ごとの野生のアマノリ養殖も注目されている[61][63]。

日本以外では、韓国、中国、台湾などでアマノリ養殖が行われている[49][64][65] (右図2i)。韓国ではスサビノリの他に、オニアマノリ、イチマツノリ、マルバアサクサノリ、ウップルイノリなどが用いられている[49][66]。また中国では、長江河口以南でハイタンアマノリ、以北でスサビノリが養殖されている[49]。
病害
上記のようなさまざまな技術開発により、21世紀には以前に比べて安定したアマノリ生産が可能になっている。しかしそれでも、寄生生物による病害や環境条件による生育阻害が起こることがある。
- 壺状菌病 (chytrid disease, Olpidiopsis disease)[5][67][68][69]
- アマノリ葉状体 (配偶体) が緑色に変色し、さらに悪化すると白化して網から脱落する。卵菌類のフクロカビモドキ属の1種 (Olpidiopsis porphyrae) の寄生によって引き起こされる。
- あかぐされ病 (red-rot disease)[5][68][70]
- アマノリ葉状体の表面に赤さび色の斑点が出来て腐敗する。卵菌類のフハイカビ属の1種 (Pythium porphyrae) の寄生によって引き起こされる。環境条件としては低塩分濃度、高水温が続いた場合に起こりやすい。干出や酸処理によって改善することがある。
- White spot disease[68][69]
- Diatom felt[68]
- アマノリ葉状体の表面が汚れ、一部は白化する。珪藻 (Melosira, Licmophora, Navicula など) が多数着生することで起こる。干出や酸処理によって改善することがある。
- Cyanobacteria felt[68]
- アマノリ葉状体の表面が汚れ、一部は孔が開く。シアノバクテリア (藍藻) が多数着生することで起こる。干出や酸処理によって改善することがある。
- Greening-spot disease[68]
- アマノリ葉状体に緑色の縞が生じ、腐敗して孔が開く。複数種の細菌 (Flabobacterium sp., Pseudoalteromonas sp., Vibrio sp. など) が原因となる。
- スミノリ病[71]
- 外見的な症状はほとんどないが、アマノリ葉状体を淡水に浸すと原形質が漏出する。このため板海苔に漉く際に細胞が壊れ、艶のない板海苔になってしまう。複数種の細菌 (Gaetbulibacter sp. など) が原因となるとともに、アマノリの生理的状態が悪化した際に発病すると考えられている。
- しろぐされ病[5]
- アマノリ葉状体が白化し、2-3週間で壊死する。干出不足や光量不足による生理的障害と考えられている。
- 芽いたみ病[5]
- アマノリ葉状体の幼芽期から幼葉期にかけて成長が止まり脱落する。原因は不明だが、何らかの生理的障害と考えられている。
- 色落ち[5][72]
岩海苔
養殖ではない天然のアマノリ類を採取して食用とする場合、日本ではこれを岩海苔 (いわのり) とよぶ[57]。地域によって種は異なり、ウップルイノリ、オニアマノリ、マルバアマノリ、ツクシアマノリなどが岩海苔となる。また養殖種として最も多く利用されるスサビノリであっても、野生の個体は岩海苔となる。一方でウップルイノリなど岩海苔として一般的な種であっても、養殖した場合は岩海苔と称することはできない (景品表示法第5条 (第4条第1項) 第1号 (優良誤認))。
岩海苔の採取が盛んな地域では、その増殖を目的としてコンクリート製のノリ礁を造成したり、雑藻駆除が行われることもある[27]。
韓国では、オニアマノリ、イチマツノリ、マルバアマノリ、ウップルイノリのようなスサビノリ以外のアマノリが養殖されることも多い。これらのアマノリは「岩海苔」とよばれ、スサビノリよりも高価に取引される[49][66]。
食用

アマノリは生のまま食用とされることもある (生ノリ)[57]。しかし多くは水洗されて細かく裁断され、これを一定量ずつ簀上に抄き、乾燥させることで板海苔とする (右図)。現在では、板海苔はふつう全自動乾海苔製造装置を用いてつくられる (1時間に7,000枚が製造可能)[27][57]。日本では板海苔の単位に規格があり、1枚は 21 x 19 cm、これが10枚で1帖 (じょう)、10帖で1束として結束し、36束で1箱として出荷される[5]。生産者が出荷するこの状態の板海苔は乾海苔 (ほしのり、直接消費者に流通する乾海苔は「干海苔」とも表記される) とよばれる[57]。流通・加工業者がこれを焼加工して焼海苔 (焼き海苔) としたり、焼加工し更に味付加工して味付海苔 (味付け海苔) とする[57]。また生ノリをそのまま乾燥させた製品もあり、ばら干海苔とよばれる[57]。アマノリの抽出物を健康食品に利用することもある[27]。中国では、アマノリを刻むことなく集めて円形に成形、乾燥させて圓紫菜 (円紫菜) とすることもある[73]。
日本においてアマノリを用いた最も身近な料理として、おにぎりや寿司がある (下図)。特に寿司が国際的な料理になったため、海苔 (アマノリ) も国際的に知られるようになった (英語でもnori とよばれる)。他にも海苔弁当や茶漬け、ざるそば、海苔煎餅、磯辺餅など、さまざまな食品に利用されている (下図)。佃煮とすることもあるが、市販されている"海苔の佃煮"の多くは紅藻のアマノリではなく緑藻のヒトエグサを原料としている[74]。

.jpg.webp)
.jpg.webp)


韓国では、調味料で味付けをしてごま油で炙った韓国海苔がよく知られている (下図)[75][76]。他にもビビンバに振りかけたり、ご飯と野菜、キムチ、ハム、チーズなどを巻いたキムパプなどがある (下図)。中国や台湾、タイなどの沿岸域では、アマノリをスープにし、これに白身魚のすり身などを入れることが多い[77] (下図)。



_on_Toast.jpg.webp)
ブリティッシュコロンビア州 (カナダ) の先住民は採取したアマノリを乾燥し、サーモンの付け合わせとしたり粉末にして食べる[75]。またハワイではアマノリ (現地名:limu luau) が珍味とされる[75][78]。フィリピンではアマノリを円盤状の形に乾燥させて "gamet" とし、これを肉や魚の付け合わせとしたり、サラダに用いる[75]。
イギリス (特にウェールズ) ではアマノリ類の Porphyra umbilicalis (現地名:laver, sloak) を採取し、長時間煮てペースト状にする[79]。これを付け合わせにしたり、オートミールを混ぜて揚げたりする。調理したものは laverbread ともよばれる (上図)。
成分
| 100 gあたりの栄養価 | |
|---|---|
| エネルギー | 787 kJ (188 kcal) |
|
44.3 g | |
| 食物繊維 | 36.0 g |
|
3.7 g | |
| 飽和脂肪酸 | 0.55 g |
| 一価不飽和 | 0.20 g |
| 多価不飽和 | 1.39 g |
|
41.4 g | |
| ビタミン | |
| ビタミンA相当量 |
(288%) 2300 µg(231%) 25000 µg |
| チアミン (B1) |
(60%) 0.69 mg |
| リボフラビン (B2) |
(194%) 2.33 mg |
| ナイアシン (B3) |
(78%) 11.7 mg |
| パントテン酸 (B5) |
(24%) 1.18 mg |
| ビタミンB6 |
(45%) 0.59 mg |
| 葉酸 (B9) |
(475%) 1900 µg |
| ビタミンB12 |
(2400%) 57.6 µg |
| ビタミンC |
(253%) 210 mg |
| ビタミンE |
(31%) 4.6 mg |
| ビタミンK |
(371%) 390 µg |
| ミネラル | |
| ナトリウム |
(35%) 530 mg |
| カリウム |
(51%) 2400 mg |
| カルシウム |
(28%) 280 mg |
| マグネシウム |
(85%) 300 mg |
| リン |
(100%) 700 mg |
| 鉄分 |
(88%) 11.4 mg |
| 亜鉛 |
(38%) 3.6 mg |
| 銅 |
(28%) 0.55 mg |
| セレン |
(13%) 9 µg |
| 他の成分 | |
| 水分 | 2.3 g |
| コレステロール | 22 mg |
| ヨウ素 | 2100 μg |
| ビオチン (B7) | 46.9 μg |
| |
| %はアメリカ合衆国における 成人栄養摂取目標 (RDI) の割合。 | |
アマノリはふつう板海苔の形で流通しており、板海苔は加工度が高い食品である[57]。タンパク質や食物繊維、ミネラル (鉄、亜鉛、カリウム、カルシウム、マンガン、マグネシウム) の割合が多い[57] (右表)。ビタミンはA、B1 (チアミン)、B2 (リボフラビン)、B9 (葉酸)、B12、C (アスコルビン酸)、脂肪酸はEPA、他にタウリンやβ-カロテンを多く含む[57] (右表、下表)。
アマノリ葉状体の細胞壁の繊維性多糖はキシランやマンナンであるが、基質となる粘質多糖としてポルフィランを含む。ヒトは自身ではポルフィランを分解できないためふつうこの多糖は食物繊維となるが、一部の日本人の腸内細菌はポルフィラン分解酵素をもつことが知られている[81]。このポルフィラン分解酵素はもともと腸内細菌には存在せず、海生細菌からの遺伝子水平伝播によるものと考えられている[81]。日本人は古くから海藻を食用としていたため、海藻などに付随していた海生細菌が消化管に入り、その分解酵素遺伝子が水平伝播によって腸内細菌に取り込まれ、この腸内細菌が一部の日本人に受け継がれてきたと考えられている[81]。
アマノリの呈味成分には遊離アミノ酸、遊離糖、糖アルコール、核酸、有機酸などがあり、特に遊離アミノ酸であるグルタミン酸、アラニン、アスパラギン酸が重要であると考えられている[57] (下表)。他にイノシン酸やグアニル酸も報告されている[57]。
焼海苔の香りは、アルデヒド (2-メチルブタナール、3-メチルブタナール)、含硫黄化合物 (3-メチルチオフェン、2,5-ジメチルチオフェン)、窒素を含む複素環芳香族化合物 (2-メチルピラジン、2,5-ジメチルピラジン、2-エチル-3,6-ジメチルピラジン、3-メチルピロール) が主成分となる[57]。
| タウリン | 2,081 | アスパラギン酸 | 142 |
| スレオニン | 24 | セリン | 23 |
| グルタミン酸 | 567 | グルタミン | 42 |
| サルコシン | 19 | プロリン | 72 |
| グリシン | 38 | アラニン | 1,254 |
| シトルリン | 75 | α-アミノ酪酸 | 21 |
| バリン | 31 | メチオニン | 6 |
| イソロイシン | 17 | ロイシン | 23 |
| チロシン | 12 | フェニルアラニン | 14 |
| β-アラニン | 19 | β-アミノイソ酪酸 | 9 |
| γ-アミノ酪酸 | 26 | ヒスチジン | 5 |
| トリプトファン | 4 | オルニチン | 1 |
| リジン | 9 | アルギニン | 12 |
分類
分類の変遷
アマノリは紅藻のウシケノリ綱、ウシケノリ目、ウシケノリ科に属する。2011年まではふつうポルフィラ属 (Porphyra; アマノリ属とよばれていた) にまとめられていた[2]。また同じウシケノリ科に属し、多列糸状の体をもつ種はウシケノリ属 (Bangia) にまとめられていた。しかし詳細な分子系統学的研究が行われることによって、この意味でのアマノリ属、ウシケノリ属は単系統群ではなく、系統的には両者が混ざった状態であることが示された[83] (下図)。そのため古典的な意味でのアマノリ属は解体され、2020年現在では14属に分けられている[83][84]。ただしこの分類は主に分子形質に基づいており、分子情報が得られていない種については分類すべき属が明らかになっていない。このような種は暫定的にポルフィラ属に残されている[2]。
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ウシケノリ綱内の系統仮説の一例[83][84][85].● = アマノリ類 (膜状の体をもつ属)。 二重線は非単系統群であることを示す。 アマノリ類は非単系統群であることを示している。 |
また2011年以前には、和名ではポルフィラ属 (Porphyra) をアマノリ属とよんでいた。しかしアサクサノリやスサビノリが分類される属が Pyropia に変更され、さらに Neopyropia と変更されるのに伴い、アマノリ属に充てる属も変更されている[2]。
14属約160種が知られており、日本からは9属約30種が報告されている[86][87]。ただし日本産のうちポルフィラ属6種は分子系統学的情報がないため暫定的にポルフィラ属に残されている種であり、将来的にはポルフィラ属以外の属に移される可能性が高い[2]。
食用種
アマノリ類は、食用とされるまたは食用とされていた種を多く含む[88]。以下にその例を示す。
- スサビノリ (尻沢辺海苔、学名:Neopyropia yezoensis)[7][89][90] (→詳細は「スサビノリ」を参照)
- 2020年現在、日本における主要な食用・養殖種である。葉状体は卵形から長楕円形。大きさはふつう 5-20 x 2–8 cm。赤褐色で基部がふつう緑色がかる。1細胞層で厚さは25-53 μm。縁辺に顕微鏡的な鋸歯はない。雌雄同株 (雌雄生殖細胞は混在) だが、雌雄異株の個体も存在。精子嚢分裂様式は64-128 (2?4 x 4 x 8)、果胞子分裂様式は16 (2 x 2 x 4)。原胞子による無性生殖あり。品集のナラワスサビノリ (Neopyropia yezoennsis f. narawaensis) が養殖に広く用いられている。北海道から本州北部、朝鮮半島に分布。外海に面した岩礁帯の潮間帯下部から上部に生育。
- アサクサノリ (浅草海苔、学名:Neopyropia tenera)[7][23][90] (→詳細は「アサクサノリ」を参照)
- スサビノリが利用される以前は、主な食用・養殖種であった。現在では野生個体は絶滅危惧I類に指定されている[91]。葉状体は長楕円形から線状披針形。大きさはふつう 7-24 x 2–5 cm。淡褐色から緑色がかった赤褐色、基部はときに青緑色がかる。1細胞層で厚さは14-35 μm。縁辺に顕微鏡的な鋸歯はない。雌雄同株 (雌雄生殖細胞は混在) または雄個体が存在。精子嚢分裂様式はふつう64 (4 x 4 x 4)、果胞子分裂様式は8 (2 x 2 x 2)。原胞子による無性生殖あり。北海道南部から九州、朝鮮半島に分布。河口や干潟に生育。
- マルバアサクサノリ (丸葉浅草海苔、学名:Neopyropia kuniedae)[7][28]
- 古くは養殖されていたが、2020年現在は絶滅危惧I類に指定されている[91]。葉状体は円形から腎臓形。大きさは 10 x 15 cm ほどまで。やや赤味のある褐色。縁辺は全縁、皺状に波打ったり巻き込んでおり、ときに基部まで裂けている。1細胞層で厚さは28-35 μm。縁辺に顕微鏡的な鋸歯はない。雌雄同株 (雌雄生殖細胞は混在)。精子嚢分裂様式は32 (4 x 2 x 4)、果胞子分裂様式は4 (2 x 1 x 2)。原胞子による無性生殖あり。東北太平洋岸、広島湾、有明海、朝鮮半島に分布。潮間帯に生育。春から夏にも葉状体が残存することがある。
- ヤブレアマノリ (学名:Neopyropia lacerata)[7][92]
- 古くは養殖されていた。葉状体は長円形から腎臓形。大きさはふつう 5 x 5 cm ほど。赤褐色から黄褐色。縁辺は全縁、成熟すると裂ける。1細胞層で厚さは24-30 μm。縁辺に顕微鏡的な鋸歯はない。雌雄同株 (精子嚢斑は縦長)。精子嚢分裂様式は64 (4 x 4 x 4)、果胞子分裂様式は4 (1 x 2 x 2)。原胞子による無性生殖あり。本州太平洋岸中部、九州西岸、朝鮮半島に分布。外海に面した潮間帯上部に生育。ときに他の海藻に着生する。
- ソメワケアマノリ (学名:Neopyropia katadae)[7][25]
- 古くは養殖されていたが、2020年現在は準絶滅危惧に指定されている[91]。葉状体は卵形から披針形、ときに勾玉状。大きさはふつう 5-15 x 2–6 cm ほど (大きいものは 20 x 16 cm に達する)。やや赤みがかった暗褐色。縁辺は全縁、成熟すると裂ける。1細胞層で厚さは20-30 μm。縁辺は全縁でやや波打ち、顕微鏡的な鋸歯はない。雌雄同株で雄の部分と雌の部分が左右に分かれている。精子嚢分裂様式は64 (4 x 4 x 4)、果胞子分裂様式は16 (2 x 2 x 4)。北海道から本州太平洋岸中部、および九州、朝鮮半島に分布。河口の低潮線付近や干潟に生育し、しばしばオゴノリやウツロムカデ (ともに紅藻) に着生する。
- カイガラアマノリ (貝殻甘海苔、学名:Neopyropia tenuipedalis)[7][20][61]
- 絶滅危惧I類に指定されているが[91]、山口県では養殖も試みられている。葉状体は長円形~披針形。大きさはふつう 8-30 x 2–9 cm (ときに 60 x 10 cm になる)。鮮やかな紅色から橙紅色。1細胞層で厚さは14-42 μm。縁辺に顕微鏡的な鋸歯はない。雌雄同株 (雌雄の生殖細胞は縁辺部で混在)。精子嚢分裂様式は128 (4 x 4 x 8)、果胞子分裂様式は16 (2 x 2 x 4)。原胞子による無性生殖はない。殻胞子を欠き、貝殻中の糸状体から直接葉状体が形成される (そのため葉状体は貝殻から生じている)。東京湾から瀬戸内海に分布。潮下帯に生育。
- ウップルイノリ (十六島海苔、学名:Pyropia pseudolinearis)[7][22]
- マルバアマノリ (丸葉甘海苔、学名:Phycocalidia suborbiculata)[7][93]
- 野生個体が岩海苔として利用されることがある。葉状体は円形~腎臓形。大きさは 10 x 10 cm 程度。赤褐色。1細胞層で厚さは30-48 μm。縁辺に顕微鏡的な鋸歯がある。雌雄同株 (雌雄の生殖細胞は混在)。精子嚢分裂様式は64 (4 x 4 x 4)、果胞子分裂様式は32 (2 x 4 x 4)。原胞子による無性生殖を行う。太平洋、大西洋、インド洋の沿岸に広く分布し、日本では北海道南部から南西諸島まで見られる。
- ツクシアマノリ (筑紫甘海苔、学名:Phycocalidia acanthophora)[7][94]
- 野生個体が岩海苔として利用されることがある (和歌山県など)。葉状体は円形~腎臓形、ときに縮れて団塊状。大きさは 4 x 4 cm 程度。1細胞層で厚さは45-50 μm。縁辺に顕微鏡的な鋸歯がある。雌雄同株 (雌雄の生殖細胞は混在)。精子嚢分裂様式は128 (4 x 4 x 8)、果胞子分裂様式は16または32 (2 x 2-4 x 4)。本州太平洋岸中部から南西諸島、東南アジアに分布。潮間帯上部に生育。
- イチマツノリ (市松海苔、学名:Neoporphyra seriata)[7][95]
- 韓国では養殖されることがある。葉状体は円形~腎臓形。大きさはふつう縦横 5–12 cm ほど、ときに 25 cm に達する。濃い緑色がかった褐色。1細胞層で厚さは38-74 μm。縁辺はやや波打ち、ときに裂片を生じる。顕微鏡的な鋸歯がない。雌雄同株、精子嚢斑はほぼ正方形で市松模様を形成する。精子嚢分裂様式は128 (4 x 4 x 8)、果胞子分裂様式は16 (2 x 2 x 4)。原胞子による無性生殖はない。北海道南部、本州北部、山口県、九州、朝鮮半島、中国に分布。潮間帯に生育する。
- オニアマノリ (鬼甘海苔、学名:Neoporphyra dentata)[7][96]
- 野生個体が岩海苔として利用されることがある。葉状体は長卵形~線状披針形。大きさはふつう 10-15 x 2–4 cm、ときに 40 x 10 cm に達する。赤褐色。1細胞層で厚さは30-58 μm。縁辺に顕微鏡的な鋸歯がある。雌雄異株、ときに雌雄が上下に分かれて形成される雌雄同株の個体がある。精子嚢分裂様式は128 (4 x 4 x 8)、果胞子分裂様式は16 (2 x 2 x 4)。原胞子による無性生殖はない。北海道南部から九州、朝鮮半島に分布。波の強い潮間帯上部に生育する。
- ハイタンアマノリ (ハイタネンシス、タンシサイ、タンシンノリ、坛紫菜、壇紫菜、学名:Neoporphyra haitanensis)[97]
- 中国福建省、浙江省、広東省沿岸で主要な養殖種となっている。葉状体は長円形~長披針形。大きさはふつう 12-18 x 3–5 cm (長さ 4 m に達することがある)。暗緑紫色。1細胞層で厚さは35-50 μm。縁辺はわずかに波打ち鋸歯状、顕微鏡的な鋸歯あり。雌雄同株だが、初期には単性。原胞子による無性生殖は知られていない。中国沿岸部に分布。波の強い潮間帯に生育。
- コスジノリ (学名:Porphyra angusta)[2][7]
- 古くは養殖に用いられていたが近年では採集記録がなく、2020年現在の環境省レッドリストでは絶滅種とされる[91]。葉状体は披針形。大きさはふつう 6-10 x 1-1.5 cm。1細胞層で厚さは25-36 μm。縁辺は全縁で波打ち、顕微鏡的な鋸歯はない。雌雄異株。精子嚢分裂様式は128 (4 x 4 x 8)、果胞子分裂様式は8 (2 x 2 x 2)。受精突起は顕著に突出する。本州太平洋岸中部に分布。外海に面した潮間帯に生育。2020年現在、分子情報が得られていないため暫定的にポルフィラ属に残されている。
- ムロネアマノリ (学名:Porphyra akasakae)[2][7]
- 古くはコスジノリの名で養殖に用いられていた。葉状体は楕円形~披針形。大きさはふつう 9-24 x 1.5–9 cm。1細胞層で厚さは25-30 μm。縁辺は全縁で波打ち、顕微鏡的な鋸歯はない。雌雄異株。精子嚢分裂様式は128 (4 x 4 x 8)、果胞子分裂様式は8-16 (2 x 2 x 2-4)。受精突起はあまり突出しない。原胞子による無性生殖が知られている。本州太平洋岸北部に分布。2020年現在、分子情報が得られていないため暫定的にポルフィラ属に残されている。
- laver (学名:Porphyra umbilicalis)[76][98] (→詳細は「Porphyra umbilicalis」を参照)
脚注
出典
- 宮下 章 (1974). “神仙菜 (アマノリ) 紫菜 (ムラサキノリ)”. 海藻. 法政大学出版局. pp. 62-64. ISBN 978-4-588-20111-0
- 有賀 祐勝 (2020年). “アマノリ類の学名”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- 堀 輝三 (編) (1993). 藻類の生活史集成. 褐藻・紅藻類. pp. 200-217. ISBN 978-4753640584
- 千原 光雄 (1997). 藻類多様性の生物学. 内田老鶴圃. pp. 55-59. ISBN 978-4753640607
- 鬼頭 釣 (監), ed (2004). わが国の水産業 のり. 社団法人 日本水産資源保護協会. pp. 1-15
- 能登谷正浩「海苔研究の現状と課題」『藻類』第48巻、2000年、247-252頁、NAID 10006435762。
- 吉田 忠生 (1998). 新日本海藻誌 日本海藻類総覧. 内田老鶴圃. pp. 438-453. ISBN 978-4753640492
- 有賀 祐勝 (2020年). “オオバアサクサノリ、ナラワスサビノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- Graham, J.E., Wilcox, L.W. & Graham, L.E. (2008). “Red Algae”. Algae. Benjamin Cummings. pp. 337-338. ISBN 978-0321559654
- Lee, R. E. & Fultz, S. A. (1970). “Ultrastructure of the Conchocelis stage of the marine red alga Porphyra leucosticta”. Journal of Phycology 6: 22-28. doi:10.1111/j.1529-8817.1970.tb02351.x.
- Gretz, M. R., Aronson, J. M. & Sommerfeld, M. R. (1980). “Cellulose in the cell walls of the Bangiophyceae (Rhodophyta)”. Science 207: 779-781. doi:10.1126/science.207.4432.779.
- Nelson, W. A., Brodie, J. & Guiry, M. D. (1999). “Terminology used to describe reproduction and life history stages in the genus Porphyra (Bangiales, Rhodophyta)”. Journal of Applied Phycology 11: 407-410. doi:10.1023/A:1008174307352.
- Drew, K. M. (1954). “Studies in the Bangioideae III. The Life-history of Porphyra umbilicalis (L.) Kütz. var. lociniata (Lightf.) J. Ag. A. The Conchocelis-phase in culture”. Annals of Botany 18 (2): 183-211.
- 右田 清治 & 伊藤 龍星 (1987). “培養によるタネガシマアマノリの生活史”. 長崎大学水産学部研究報告 61: 7-14.
- Wang, D., Yu, X., Xu, K., Bi, G., Cao, M., Zelzion, E., ... & Du, G. (2020). “Pyropia yezoensis genome reveals diverse mechanisms of carbon acquisition in the intertidal environment”. Nature Communications 11 (1): 4028. doi:10.1038/s41467-020-17689-1.
- Jiang, H., Gao, K. & Helbling, E. W. (2009). “The conchocelis of Porphyra haitanensis (Rhodophyta) is protected from harmful UV radiation by the covering calcareous matrix”. Journal of Phycology 45 (6): 1270-1277. doi:10.1111/j.1529-8817.2009.00755.x.
- Pueschel, C. M. & Cole, K. M. (1982). “Rhodophycean pit plugs: an ultrastructural survey with taxonomic implications”. American Journal of Botany 69: 703-720. doi:10.1002/j.1537-2197.1982.tb13310.x.
- Takaichi, S., Yokoyama, A., Mochimaru, M., Uchida, H. & Murakami, A. (2016). “Carotenogenesis diversification in phylogenetic lineages of Rhodophyta”. Journal of Phycology 52: 329-338. doi:10.1111/jpy.12411./
- 広田望「海藻の色」『調理科学』第13巻第4号、日本調理科学会、1980年、256-263頁、doi:10.11402/cookeryscience1968.13.4_256、ISSN 0910-5360、NAID 110001802105。
- 有賀 祐勝 (2020年). “カイガラアマノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- 有賀 祐勝 (2020年). “ベニタサ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年8月8日閲覧。
- 有賀 祐勝 (2013年). “ウップルイノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- 有賀 祐勝 (2020年). “アサクサノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- 有賀 祐勝 (2020年). “カヤベノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年8月8日閲覧。
- 有賀 祐勝 (2020年). “ソメワケアマノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年8月8日閲覧。
- 有賀 祐勝 (2020年). “ベンテンアマノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年8月8日閲覧。
- 菊池 則雄 (2012). “アマノリ”. In 渡邉 信 (監). 藻類ハンドブック. エヌ・ティー・エス. pp. 611-616. ISBN 978-4864690027
- 有賀 祐勝 (2020年). “マルバアサクサノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- 有賀 祐勝 (2020年). “チシマクロノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年8月8日閲覧。
- 宮下 章 (1974). “海藻の初見”. 海藻. 法政大学出版局. pp. 49-53. ISBN 978-4-588-20111-0
- “海苔の日”. のりホームページ. 全国海苔貝類漁業協同組合連合会. 2020年8月7日閲覧。
- “2月6日 海苔の日”. なるほど統計学園. 総務省 統計局. 2020年8月7日閲覧。
- 宮下章 (1974). “貢納品となる”. 海藻. 法政大学出版局. pp. 72-81. ISBN 978-4-588-20111-0
- 演田仁「出雲園・十六島 (うつぶるい) とウップルイノリ」(PDF)『藻類』第55巻、日本藻類学会、2007年、121-122頁。
- “木簡庫”. 奈良文化財研究所. 2020年8月24日閲覧。
- 宮下 章 (1974). “神話の中の海藻”. 海藻. 法政大学出版局. pp. 18-23. ISBN 978-4-588-20111-0
- 宮下 章 (1974). “国別特産”. 海藻. 法政大学出版局. pp. 243-251. ISBN 978-4-588-20111-0
- 宮下 章 (1974). “貴族と伏庵の民の藻食”. 海藻. 法政大学出版局. pp. 81-86. ISBN 978-4-588-20111-0
- 宮下 章 (1974). “海藻の全国的交流”. 海藻. 法政大学出版局. pp. 87-102. ISBN 978-4-588-20111-0
- 宮下 章 (1974). “中世の海藻料理”. 海藻. 法政大学出版局. pp. 103-118. ISBN 978-4-588-20111-0
- 宮下 章 (1974). “ノリを売る店”. 海藻. 法政大学出版局. pp. 171-176. ISBN 978-4-588-20111-0
- 宮下 章 (1974). “浅草海苔の誕生”. 海藻. 法政大学出版局. pp. 164-171. ISBN 978-4-588-20111-0
- 殖田三郎「養殖技術発達の沿革と現状」『水産増殖』第4巻第4号、日本水産増殖学会、1957年、8-10頁、doi:10.11233/aquaculturesci1953.4.4_8、ISSN 0371-4217、NAID 130003714610。
- 宮下 章 (1974). “ノリ産地の拡大”. 海藻. 法政大学出版局. pp. 180-190. ISBN 978-4-588-20111-0
- 金子政之助「ノリ種苗移殖の意義と方法」『水産増殖』第4巻、1957年、14-17頁、doi:10.11233/aquaculturesci1953.4.4_14。
- Drew, K.M. (1949). “Conchocelis-phase in the life-history of Porphyra umbilicalis (L.) Kütz”. Nature 164: 748-749. doi:10.1038/1647.
- 杉田 隆治「人工種付の現況と将来」『水産増殖』第4巻、1957年、52-54頁、doi:10.11233/aquaculturesci1953.4.4_52。
- 倉掛 武雄「人工種付の現況と将来」『水産増殖』第4巻、1957年、60-64頁、doi:10.11233/aquaculturesci1953.4.4_60。
- 石渡 誠之「海苔業界の現況:日本・韓国・中国」『藻類』第53巻、2005年、49-55頁。
- 須藤 俊造「東京湾を主とした養殖ノリの種類」『水産増殖』第4巻、1957年、28-30頁、doi:10.11233/aquaculturesci1953.4.4_28。
- 田中 剛「ノリの種類」『水産増殖』第4巻、1957年、18-20頁、doi:10.11233/aquaculturesci1953.4.4_18。
- 黒木 宗尚「養殖ノリの種類」『水産増殖』第4巻、1957年、21-28頁、doi:10.11233/aquaculturesci1953.4.4_21。
- 木村 五郎「躍進するノリ養殖業の展望」『水産増殖』第4巻、1957年、1-3頁、doi:10.11233/aquaculturesci1953.4.4_1。
- 齋藤 壽典 (2012年). “海苔増殖の温故知新”. 海苔百景. 一般財団法人 海苔増殖振興会. 2020年8月8日閲覧。
- “年度別の産地別生産量”. 一般財団法人 海苔増殖振興会. 2020年9月16日閲覧。
- 村山健二, 石川幹子「東京湾における地先海面の共同利用の歴史的変遷に関する研究 : 大森の海苔養殖を事例として」『都市計画. 別冊都市計画論文集』第45巻第3号、日本都市計画学会、2010年10月、403-408頁、doi:10.11361/journalcpij.45.3.403、ISSN 09160647、NAID 10028989100。
- 海苔の成分の効用と利用. 一般財団法人海苔増殖振興会. (2019). pp. 1-30
- “海面漁業生産統計調査”. e-Stat. 2020年7月23日閲覧。
- “漁業生産のうち海苔養殖業の占める割合”. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- 藤井 弘治 (2020年). “のり産業界の現状”. 産地を追って. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- 阿部 真比古「水産としての海藻」『藻類』第64巻、2016年、107-111頁。
- 阿部真比古, 小林正裕, 玉城泉也, 藤吉栄次, 菊地則雄「ATP6遺伝子に関連したミトコンドリアDNA部分塩基配列を用いた変種オオバアサクサノリ Porphyra tenera var. tamatsuensis の判別について(予報)」『水産増殖』第56巻、2008年、497-503頁、doi:10.11233/aquaculturesci.56.497、NAID 10026140058。
- “国内最高値つけた「幻のノリ」、全滅から復活の兆し”. 朝日新聞デジタル (2020年2月4日). 2020年8月11日閲覧。
- McHugh, D. J. (2003年). “8. SEAWEEDS USED AS HUMAN FOOD”. A guide to the seaweed industry. FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS. 2020年8月3日閲覧。
- Buschmann, A. H., Camus, C., Infante, J., Neori, A., Israel, Á., Hernández-González, M. C., ... & Critchley, A. T. (2017). “Seaweed production: overview of the global state of exploitation, farming and emerging research activity”. European Journal of Phycology 52 (4): 391-406. doi:10.1080/09670262.2017.1365175.
- 有賀 祐勝 (2015年). “韓国の海苔・中国の海苔”. 海苔百景. 一般財団法人 海苔増殖振興会. 2020年8月8日閲覧。
- 本多 大輔「壺状菌の分類と漁場における探索」『日本水産学会誌』第75巻、2009年、908-909頁、doi:10.2331/suisan.75.908。
- Kim, G. H., Moon, K. H., Kim, J. Y., Shim, J. & Klochkova, T. A. (2014). “A revaluation of algal diseases in Korean Pyropia (Porphyra) sea farms and their economic impact”. Algae 29 (4): 249-265. doi:10.4490/algae.2014.29.4.249.
- Guan, X., Li, J., Zhang, Z., Li, F., Yang, R., Jiang, P. & Qin, S. (2013). “Characterizing the microbial culprit of white spot disease of the conchocelis stage of Porphyra yezoensis (Bangiales, Rhodophyta)”. Journal of Applied Phycology 25 (5): 1341-1348. doi:10.1007/s10811-013-9976-8.
- 東條 元昭「ノリアカグサレ菌の分類学的研究」『日本水産学会誌』第75巻、2009年、910-911頁、doi:10.2331/suisan.75.910。
- 三根 崇幸「海苔スミノリ病の発症要因解析と防除法開発に関する研究」『佐賀県有明水産振興センター研究報告』第25巻、2011年、41―76、NAID 80022164437。
- 堀豊, 望月松寿, 島本信夫「播磨灘北部海域におけるノリ色落ちと漁場環境の変遷」『水産海洋研究』第72巻、2008年、107-112頁、NAID 10024766911。
- 有賀 祐勝 (2019年). “大連の海鮮・干物店で見た海苔・昆布・若布”. 海藻百景 リレーエッセイ. 一般財団法人 海苔増殖振興会. 2020年8月10日閲覧。
- 天野 秀臣 (2018年). “ヒトエグサの思い出”. 海藻百景 リレーエッセイ. 一般財団法人 海苔増殖振興会. 2020年8月10日閲覧。
- Robin Harford (2020年). “Laver Seaweed - A Foraging Guide to Its Food, Medicine and Other Uses”. A Wild Food Foraging Guide to the Edible and Medicinal Wild Plants of Britain. 2020年8月3日閲覧。
- カオリ・オコナー (著), 龍 和子 (訳) (2018). “朝鮮半島の海藻”. 海藻の歴史. 原書房. pp. 91-98. ISBN 978-4562054138
- 大野 正夫「世界の海藻食」『日本食育学会誌』第14巻、2020年、3-7頁、NAID 40022147666。
- Reed, M. (1907). Economic Seaweeds of Hawaii and Their Food Value. Washington, US Government Printing Office. pp. 64
- カオリ・オコナー (著), 龍 和子 (訳) (2018). “イギリス諸島と北欧の海藻”. 海藻の歴史. 原書房. pp. 131-163. ISBN 978-4562054138
- 文部科学省 「日本食品標準成分表2015年版(七訂)」
- Hehemann, J. H., Correc, G., Barbeyron, T., Helbert, W., Czjzek, M. & Michel, G. (2010). “Transfer of carbohydrate-active enzymes from marine bacteria to Japanese gut microbiota”. Nature 464: 908-915. doi:10.1038/nature08937.
- 鈴木 雅子 (1996). “乾のり製造工程における遊離アミノ酸の変化”. 千葉県水産試験場研究報告 54: 53-55. NAID 40004026389.
- Sutherland, J. E., Lindstrom, S. C., Nelson, W. A., Brodie, J., Lynch, M. D., Hwang, M. S., ... & Farr, T. (2011). “A new look at an ancient order: generic revision of the Bangiales (Rhodophyta)”. Journal of Phycology 47 (5): 1131-1151. doi:10.1111/j.1529-8817.2011.01052.x.
- Yang, L. E., Deng, Y. Y., Xu, G. P., Russell, S., Lu, Q. Q. & Brodie, J. (2020). “Redefining Pyropia (Bangiales, Rhodophyta): four new genera, resurrection of Porphyrella and description of Calidia pseudolobata sp. nov. from China”. Journal of Phycology. doi:10.1111/jpy.12992.
- Sánchez, N., Vergés, A., Peteiro, C., Sutherland, J. E. & Brodie, J. (2014). “Diversity of bladed Bangiales (Rhodophyta) in western Mediterranean: recognition of the genus Themis and descriptions of T. ballesterosii sp. nov., T. iberica sp. nov., and Pyropia parva sp. nov”. Journal of Phycology 50: 908-929. doi:10.1111/jpy.12223.
- Guiry, M.D. & Guiry, G.M.. “AlgaeBase”. World-wide electronic publication, Nat. Univ. Ireland, Galway. 2020年8月10日閲覧。
- 鈴木 雅大 (2020年7月11日). “ウシケノリ目”. 日本産海藻リスト. 2020年8月10日閲覧。
- 有賀 祐勝 (2016年). “スサビノリの「スサビ」の由来”. 海藻百景 リレーエッセイ. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- 有賀 祐勝 (2020年). “スサビノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- 菊地 則雄, 藤吉 栄次, 玉城 泉也 & 小林 正裕 (2014). “形態による判別”. In 藤吉栄次, 玉城泉也, 小林正裕 & 有瀧真人 (編). アマノリ養殖品種の特性. 水産総合研究センター 西海区水産研究所. pp. 15-23. ISBN 9784998069423
- 環境省 レッドリスト. 2020.8.12閲覧.
- 有賀 祐勝 (2020年). “ヤブレアマノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年8月10日閲覧。
- 有賀 祐勝 (2020年). “マルバアマノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- 有賀 祐勝 (2020年). “ツクシアマノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- 有賀 祐勝 (2020年). “イチマツノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- 有賀 祐勝 (2020年). “オニアマノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- 有賀 祐勝 (2020年). “ハイタンアマノリ”. 海苔の豆図鑑. 一般財団法人 海苔増殖振興会. 2020年7月23日閲覧。
- Guiry, M.D. & Guiry, G.M. (2020年2月14日). “Porphyra umbilicalis Kützing 1843”. AlgaeBase. 2020年8月12日閲覧。
外部リンク
- ウシケノリ綱 Class Bangiophyceae. 写真で見る生物の系統と分類. 生きもの好きの語る自然誌. (2020年9月27日閲覧)
- ウシケノリ目 Order BANGIALES. 日本産海藻リスト. 生きもの好きの語る自然誌. (2020年9月27日閲覧)
- 一般財団法人 海苔増殖振興会. (2020年8月10日閲覧)
- 海苔ジャーナル. 海苔産業情報センター. (2020年8月10日閲覧)
- 全国海苔貝類漁業協同組合連合会. (2020年8月10日閲覧)
- 海苔JAPAN. 海苔で健康推進委員会. (2020年8月10日閲覧)
- ノリ. せとうちネット. (2020年8月10日閲覧)
- 海苔ができるまで. 山本海苔. (2019年10月22日閲覧)
- 雑学!海苔グルメ. 岩波海苔店. (2020年8月10日閲覧)